Extracellular vesicles (EVs) act as multifunctional regulators of intercellular communication and are involved in diverse tumor phenotypes, including tumor angiogenesis, which is a highly regulated multi-step process for the formation of new blood vessels that contribute to tumor proliferation. EVs induce malignant transformation of distinct cells by transferring DNAs, proteins, lipids, and RNAs, including noncoding RNAs (ncRNAs). However, the functional relevance of EV-derived ncRNAs in tumor angiogenesis remains to be elucidated.
1. Characteristics of Extracellular Vesicles and ncRNAs
Extracellular vesicles (EVs) are extracellular structures enclosed in a lipid bilayer that can be secreted by almost all known cell types
[1]. Several studies have indicated that EVs play an essential role in intercellular communication between tumor cells and the tumor microenvironment (TME). EVs are highly heterogeneous; therefore, their characterization and classification are crucial for further research to avoid generating inconclusive results. Based on their size, biogenesis, and release pathways, EVs can be broadly divided into three main subtypes: exosomes, microvesicles (MVs), and apoptotic bodies. Exosomes, also called small EVs, possess a diameter ranging from 30 to 150 nm and are derived from multivesicular endosomal pathways, which are formed by inward budding of the endosomal membrane in a process that sequesters particular proteins and lipids
[2]. On the contrary, MVs are generated by regulated outward budding of the plasma membrane
[3]. The mechanisms of exosomal biogenesis involve multiple factors, and the most well-known regulator is endosomal sorting complex required for transport (ESCRT)
[3]. Exosomal biogenesis involves inward budding of the plasma membrane to form endosomes, leading to production of multivesicular bodies (MVBs), fusion of MVBs with the plasma membrane, and release of exosomes into the extracellular space. A core component of this mechanism is the ESCRT machinery, which consists of four protein complexes and auxiliary proteins that bind to future exosome cargoes and form intraluminal vesicles that incorporate those cargoes
[4][5]. Several studies have found that exosomes can be formed despite the depletion of the ESCRT complex, which reveals an ESCRT-independent approach
[6]. ESCRT-independent exosomal biogenesis is regulated by sphingolipid ceramide, which is produced from the hydrolysis of sphingomyelin by neutral sphingomyelinase 2
[2]. The contents of EVs include various nucleic acids, lipids, and proteins
[7]. ncRNAs carried by EVs can regulate various physiological and pathological processes through multiple mechanisms (
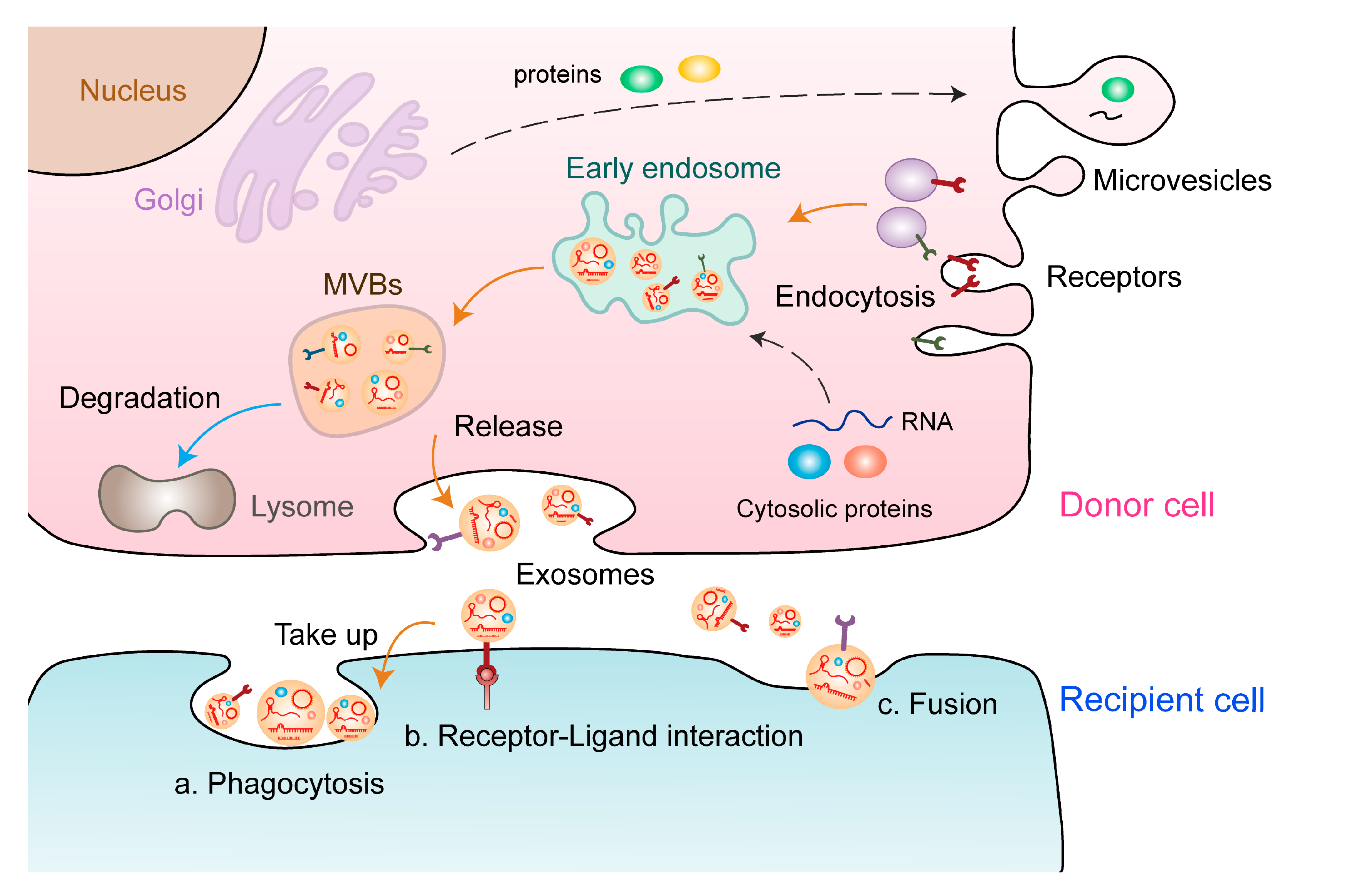
Figure 1).
Figure 1. Formation and release of EVs. The formation of EVs involves the following processes: Proteins are transported from the Golgi apparatus or internalised from the cell surface, and nucleic acids are endocytosed and transferred to early endosomes. Early endosomes gradually mature into late endosomes and MVBs, some of which are degraded, whereas others are secreted as exosomes. These exosomes carry multiple biological components, including proteins, lipids, and nucleic acids (e.g., ncRNAs), which are delivered to the recipient cell through different ways: a. phagocytosis, b. receptor–ligand interaction, and c. direct fusion.
In the past few decades, the development of high-throughput sequencing technology has indicated that the transcription rate of the human genome generally exceeds 70%. However, <2% of the transcript is translated into proteins; most human transcriptomes are ncRNAs. Emerging studies have shown that despite being ‘transcriptional junk’, ncRNAs, such as miRNAs, piRNAs, circRNAs, snoRNAs, and the attractive lncRNAs, play a versatile role in manipulating gene expression
[8]. miRNAs, a type of endogenous small RNA with a length of 20–24 nucleotides (nt), have many essential adjustable functions in cells. By complementarily binding to the 3’-untranslated region (UTR) of targeted mRNAs, miRNAs act as regulators of gene expression, thereby inhibiting post-transcriptional gene expression
[9][10][11]. Recent studies have reported that miRNAs are selectively sorted into EVs and participate in intercellular communication in the TME
[12]. In addition, EV-derived miRNAs in biofluids can be used as ideal biomarkers for various types of tumors due to their easy accessibility, high abundance, and good stability
[13]. piRNAs, a type of small RNA with a length of 21–35 nt, specifically interact with PIWI protein to perform multifaceted functions in germline development and somatic tissues
[14][15][16].
In addition to small RNAs, large ncRNAs also participate in gene regulation in various biological processes. lncRNAs, collectively referred to as transcripts with more than 200 nt, have limited potential to encode proteins
[17]. They perform their functions through multiple molecular and cellular mechanisms, such as interacting with epigenetic factors or TFs to modulate gene transcription, sequestering miRNAs, interacting with proteins, and encoding functional small peptides
[18][19][20][21]. In addition, lncRNAs can also be selectively sorted into EVs and participate in cell-to-cell communication in the TME
[22]. circRNAs, generated by a particular form of alternative splicing called back-splicing, regulate gene transcription and translation by interacting with DNAs, RNAs, and proteins
[23]. Emerging studies have indicated that circRNAs participate in multiple physiological and pathological processes, including tumor angiogenesis
[24][25][26].
2. EV-Derived ncRNAs: New Players in Tumor Angiogenesis
Angiogenesis is a multi-step process and has two types: sprouting and intussusceptive angiogenesis
[27]. Various types of cells, including endothelial cells (ECs), tumor cells, stromal cells, and immune cells, regulate angiogenesis in the blood vessels. In addition, some regulatory and signalling molecules govern angiogenesis, including growth factors (e.g., VEGF, PDGF, FGF, and EGF) and transcription factors, such as HIFs
[28][29][30]. Because angiogenesis is crucial for tumor growth and metastasis, targeting tumor-associated angiogenesis is a promising strategy for cancer treatment
[31][32][33]. Currently, anti-angiogenic therapies targeting VEGF and VEGFR are used for the treatment of various tumors
[34].
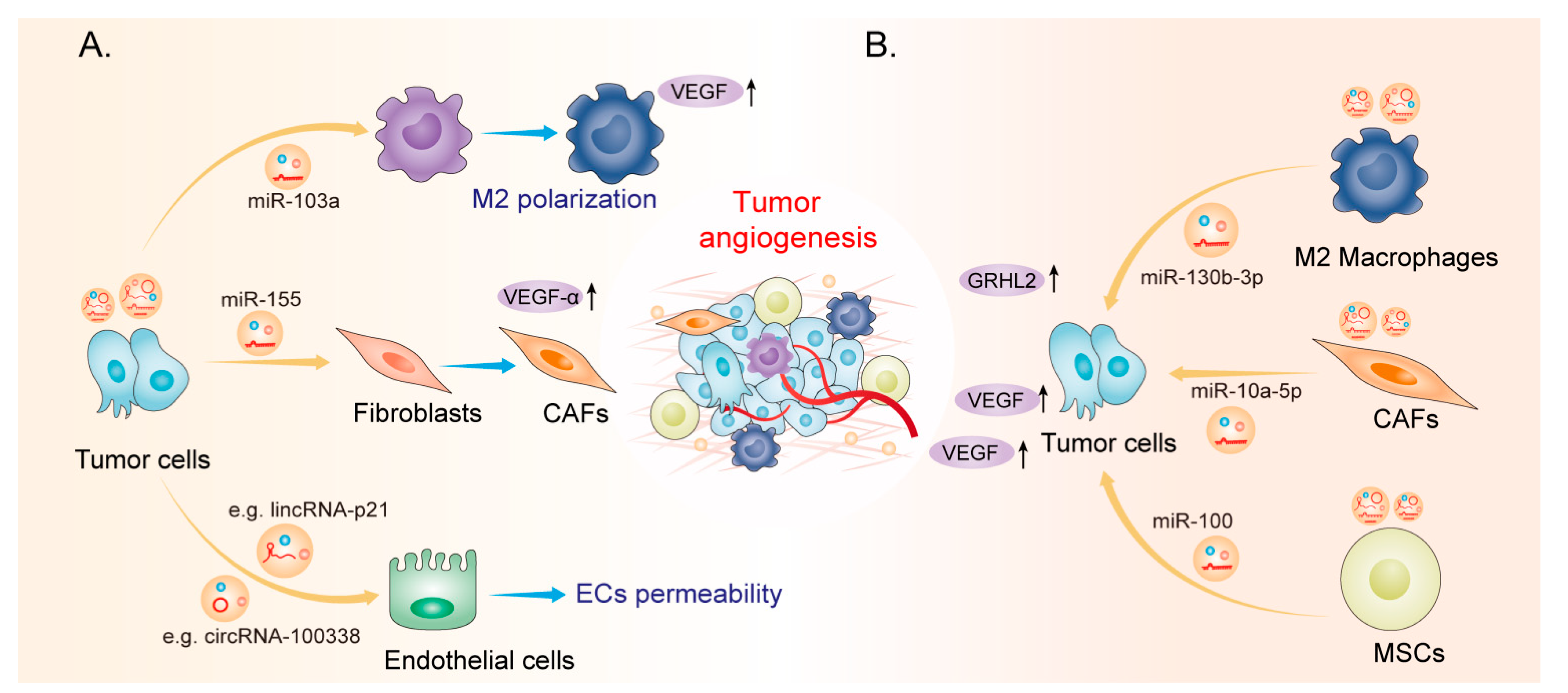
Several studies have indicated that EVs can be used as ncRNA carriers to play diverse roles in regulating tumor hallmarks, including angiogenesis. For example, in non-small cell lung cancer (NSCLC), RCAN1.4 has been identified as a target of miR-619-5p, and its suppression promotes angiogenesis
[35]. The exosomal lncRNA FAM225A upregulates NETO2 and FOXP1 expression by sponging miR-206 to accelerate oesophageal squamous cell carcinoma (ESCC) progression and angiogenesis
[36]. In addition, circSHKBP1 sponges miR-582-3p to enhance HUR expression and VEGF mRNA stability, which promotes angiogenesis and gastric tumor progression
[37]. Moreover, emerging studies have indicated that EV-derived ncRNAs can regulate tumor angiogenesis by influencing a wide variety of tumor-associated molecules. The functions and mechanisms of EV-derived ncRNAs in tumor angiogenesis are summarized in
Table 1. These studies suggest that EV-derived ncRNAs play an essential role in tumor angiogenesis. However, new technologies and animal models are required to further investigate the precise mechanisms of EV-derived ncRNAs in the regulation of tumor angiogenesis.
Table 1. The emerging roles of EV-derived ncRNAs in tumor angiogenesis.
| EV-Derived ncRNAs |
Expression |
Source Cell |
Function and Mechanism |
Tumor Type |
Reference |
|
miR-155
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis via the c-MYB/VEGF axis
|
Gastric cancer
|
[38] |
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis by inhibiting FOXO3a
|
Gastric cancer
|
[39] |
|
miR-130a
|
Upregulated
|
Tumor cell
|
Activates angiogenesis by inhibiting c-MYB
|
Gastric cancer
|
[40] |
|
miR-135b
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis by inhibiting FOXO1
|
Gastric cancer
|
[41] |
|
Upregulated
|
Tumor cell
|
Regulates the HIF/FIH signalling pathway
|
Multiple myeloma
|
[42] |
|
miR-23a
|
Upregulated
|
Tumor cell
|
Inhibits PTEN and activates the AKT pathway
|
Gastric cancer
|
[43] |
|
Upregulated
|
Tumor cell
|
Increases angiogenesis by inhibiting ZO-1
|
Lung cancer
|
[44] |
|
miR-200b-3p
|
Downregulated
|
Tumor cell
|
Enhances endothelial ERG expression
|
Hepatocellular carcinoma
|
[45] |
|
miR-25-3p
|
Upregulated
|
Tumor cell
|
Inhibits KLF2 and KLF4, thereby elevating VEGFR2 expression
|
Colorectal cancer
|
[46] |
|
miR-1229
|
Upregulated
|
Tumor cell
|
Inhibits HIPK2, thereby activating the VEGF pathway
|
Colorectal cancer
|
[47] |
|
miR-183-5p
|
Upregulated
|
Tumor cell
|
Inhibits FOXO1, thereby promoting expression of VEGFA, VEGFAR2, ANG2, PIGF, MMP-2, and MMP-9
|
Colorectal cancer
|
[48] |
|
miR-142-3p
|
Upregulated
|
Tumor cell
|
Inhibits TGFβR1
|
Lung adenocarcinoma
|
[49] |
|
miR-103a
|
Upregulated
|
Tumor cell
|
Inhibits PTEN, thereby promoting the polarization of M2 macrophages
|
Lung cancer
|
[50] |
|
miR-126
|
Upregulated
|
MSCs
|
Upregulates CD34 and CXCR4, thereby promoting expression of VEGF
|
Lung cancer
|
[51] |
|
miR-141-3p
|
Upregulated
|
Tumor cell
|
Inhibits SOCS5, thereby activating JAK/STAT3 and NF-κB signalling pathways
|
Ovarian cancer
|
[52] |
|
miR-205
|
Upregulated
|
Tumor cell
|
Regulates the PTEN/AKT pathway
|
Ovarian cancer
|
[53] |
|
miR-9
|
Downregulated
|
Tumor cell
|
Inhibits MDK, thereby regulating the PDK/AKT signalling pathway
|
Nasopharyngeal carcinoma
|
[54] |
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis by targeting COL18A1, THBS2, PTCH1, and PHD3
|
Glioma
|
[55] |
|
miR-23a
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis by inhibiting TSGA10
|
Nasopharyngeal carcinoma
|
[56] |
|
miR-210
|
Upregulated
|
Tumor cell
|
Enhances tube formation by inhibiting EFNA3
|
Leukemia
|
[57] |
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis by inhibiting SMAD4 and STAT6
|
Hepatocellular carcinoma
|
[58] |
|
miR-26a
|
Upregulated
|
Tumor cell
|
Inhibits PTEN, thereby activating the PI3K/AKT signalling pathway
|
Glioma
|
[59] |
|
miR-27a
|
Upregulated
|
Tumor cell
|
Inhibits BTG2, thereby promoting VEGF, VEGFR, MMP-2, and MMP-9 expression
|
Pancreatic cancer
|
[60] |
|
miR-155-5p
/miR-221-5p
|
Upregulated
|
M2 macrophages
|
Promotes angiogenesis by targeting E2F2
|
Pancreatic cancer
|
[61] |
|
miR-21-5p
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis by targeting TGFBI and COL4A1
|
Papillary carcinoma
|
[62] |
|
miR-100
|
- -
|
MSCs
|
Regulates the mTOR/HIF-1α signalling axis
|
Breast cancer
|
[63] |
|
miR-21
|
Upregulated
|
Tumor cell
|
Inhibits SPRY1, thereby promoting VEGF expression
|
Oesophageal squamous cell carcinoma
|
[64] |
|
Upregulated
|
Tumor cell
|
Inhibits PTEN, thereby activating PDK1/AKT signalling
|
Hepatocellular carcinoma
|
[65] |
|
miR-181b-5p
|
Upregulated
|
Tumor cell
|
Inhibits PTEN and PHLPP2, thereby activating AKT signalling
|
Oesophageal squamous cell carcinoma
|
[66] |
|
miR-9
|
Upregulated
|
Tumor cell
|
Inhibits S1P, thereby promoting VEGF expression
|
Medulloblastoma and xenoglioblastoma
|
[67] |
|
miR-10a-5p
|
Upregulated
|
CAFs
|
Inhibits TBX5, thereby activating Hedgehog signalling
|
Cervical squamous cell carcinoma
|
[68] |
|
miR-135b
|
Upregulated
|
Tumor cell
|
Enhances angiogenesis by targeting FIH-1
|
Multiple myeloma
|
[42] |
|
miR-130b-3p
|
Upregulated
|
M2 macrophages
|
Regulates the miR-130b-3p/MLL3/GRHL2 signalling cascade
|
Gastric cancer
|
[69] |
|
lncGAS5
|
Downregulated
|
Tumor cell
|
Inhibits angiogenesis by regulating the miR-29-3p/PTEN axis
|
Lung cancer
|
[70] |
|
lnc-CCAT2
|
Upregulated
|
Tumor cell
|
Promotes VEGFA and TGF-β expression
|
Glioma
|
[71] |
|
lnc-POU3F3
|
Upregulated
|
Tumor cell
|
Promotes bFGF, bFGFR, and VEGFA expression
|
Glioma
|
[72] |
|
lncRNA RAMP2-AS1
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis through the miR-2355-5p/VEGFR2 axis
|
Chondrosarcoma
|
[73] |
|
OIP5-AS1
|
Upregulated
|
Tumor cell
|
Regulates angiogenesis and autophagy through miR-153/ATG5 axis
|
Osteosarcoma
|
[74] |
|
FAM225A
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis through the miR-206/NETO2/FOXP1 axis
|
Oesophageal squamous cell carcinoma
|
[36] |
|
UCA1
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis through the miR-96-5p/AMOTL2 axis
|
Pancreatic cancer
|
[75] |
|
SNHG11
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis through the miR-324-3p/VEGFA axis
|
Pancreatic cancer
|
[76] |
|
SNHG1
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis by regulating the miR-216b-5p/JAK2 axis
|
Breast cancer
|
[77] |
|
AC073352.1
|
Upregulated
|
Tumor cell
|
Binds and stabilizes the YBX1 protein
|
Breast cancer
|
[78] |
|
MALAT1
|
Upregulated
|
Tumor cell
|
Facilitates angiogenesis and predicts poor prognosis
|
Ovarian cancer
|
[79] |
|
TUG1
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis by inhibiting caspase-3 activity
|
Cervical cancer
|
[80] |
|
LINC00161
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis and metastasis by regulating the miR-590-3p/ROCK axis
|
Hepatocellular carcinoma
|
[81] |
|
H19
|
Upregulated
|
Cancer stem cell
|
Promotes VEGF production and release in ECs
|
Liver cancer
|
[82] |
|
circSHKBP1
|
Upregulated
|
Tumor cell
|
Enhances VEGF mRNA stability by the miR-582-3p/HUR axis
|
Gastric cancer
|
[37] |
|
circRNA-100,338
|
Upregulated
|
Tumor cell
|
Facilitates HCC metastasis by enhancing invasiveness and angiogenesis
|
Hepatocellular carcinoma
|
[83] |
|
circCMTM3
|
Upregulated
|
Tumor cell
|
Promotes angiogenesis and HCC tumor growth by the miR-3619-5p/SOX9 axis
|
Hepatocellular carcinoma
|
[84] |
|
circ_0007334
|
Upregulated
|
Tumor cell
|
Accelerates CRC tumor growth and angiogenesis by the miR-577/KLF12 axis
|
Colorectal cancer
|
[85] |
|
CircFNDC3B
|
Downregulated
|
Tumor cell
|
Inhibits angiogenesis and CRC progression by the miR-937-5p/TIMP3 axis
|
Colorectal cancer
|
[86] |
|
circGLIS3
|
Upregulated
|
Tumor cell
|
Induces endothelial cell angiogenesis by promoting Ezrin T567 phosphorylation
|
Glioma
|
[87] |
|
piRNA-823
|
Upregulated
|
Tumor cell
|
Promotes VEGF and IL-6 expression
|
Multiple myeloma
|
[88] |
3. Potential Clinical Applications of EV-Derived ncRNAs in Cancers
3.1. EV-Derived ncRNAs as Promising Tumor Biomarkers
EVs that possess great potential as disease biomarkers and therapeutic carriers have attracted increasing attention
[89][90]. Biomarkers are molecules that can be used for diagnosis or prognosis. An ideal biomarker should have the following four most important characteristics: specificity, sensitivity, stability, and easy accessibility in a relatively non-invasive way. Most studies have focused on extracellular ncRNAs as potential biomarkers because they are stable and can be easily extracted from liquid biopsy, such as blood, urine, or other body fluids, using simple, sensitive, and relatively inexpensive assays. It has been reported that the quality of EV-derived ncRNAs is almost unaffected even after the samples are stored for many years because of the uniqueness of the source cell components and protection via encapsulation in the membrane
[91][92]. Therefore, EVs are stable in circulation and under various storage conditions. We conducted a comprehensive literature search of EV-derived ncRNAs from different sources and found valuable biomarkers for the diagnosis and prognosis of multiple cancers (
Figure 2A).
Figure 2. The potential clinical application of EV-derived ncRNAs in tumor angiogenesis (A) EV-derived ncRNAs can be detected from patient samples and are potential diagnostic and prognostic biomarkers. (B) A combination of targeting EV-derived ncRNAs and using conventional anti-angiogenic agents can enhance therapeutic efficacy.
Researchers are interested in investigating candidate miRNAs, lncRNAs, and circRNAs carried by EVs that may serve as biomarkers for the diagnosis, prognosis, and treatment of tumors
[93]. For example, in a study, ROC curve analysis demonstrated that plasma miR-601 and miR-760 could both be used as promising diagnostic biomarkers for advanced CRC
[94]. In a study involving patients with CRC, Ogata et al. reported that the area under the curve (AUC) was 0.953, 0.948, and 0.798. In another study, exosomal miR-23a, miR-1246, and miR-21 could distinguish CRC (all stages) from the control area
[95]. Furthermore, higher levels of plasma miR-320-EV and miR-126-EV in patients with high-risk LC can promote angiogenesis and can be significantly associated with poor overall survival
[51]. A similar study showed that serum exosomes of patients with GC were rich in lncRNA ZFAS1. In addition, the upregulation of ZFAS1 was significantly associated with tumor lymphatic metastasis and TNM staging. These studies indicate that exosomal ZFAS1 may serve as a potential prognostic biomarker for GC
[96]. Another study reported that exosomal ENSG00000258332.1 and LINC00635 could be used to differentiate patients with HCC from those with chronic hepatitis B with high specificity. Therefore, serum exosomal ENSG00000258332.1 and LINC00635, which are highly sensitive and can be obtained non-invasively, may be used as biomarkers for HCC
[97]. Similarly, recent studies have reported serum exosomal circRNAs as novel and useful tools for the non-invasive diagnosis of cancer
[98]. Exosomal circPRMT5 is highly expressed in the serum and urine of patients with bladder cancer and is closely related to tumor metastasis
[99]. In addition, certain diagnostic clinical trials are currently underway. In one such trial, exosomal lncRNAs are isolated from serum samples for the diagnosis of lung cancer (NCT03830619).
A large number of studies have focused on the diagnostic, prognostic, and therapeutic significance of EV-derived ncRNAs in different tumor types. However, the specific role of EV-derived ncRNAs in angiogenesis-related diseases remains unclear. Furthermore, almost all studies have focused on cellular experiments and EV-ncRNA-associated applications in vitro; therefore, further studies are required to validate the findings for in vivo models. In addition, the potential of EV-derived ncRNAs as biomarkers remains to be further verified in multi-center, large-scale clinical trials.
3.2 EV-Derived ncRNAs as Potential Anti-Angiogenic Therapeutic Targets
Because of their negligible antigenicity, minimal cytotoxicity, and ability to bypass endocytic pathways and phagocytosis, EVs are considered ideal natural carriers for the delivery of ncRNAs
[100]. In a study, engineered exosomes modified with DSPE–PEG2K–RGD loaded with miR-92b-3p produced synergistic anti-tumor and anti-angiogenesis effects with apatinib in nude mice models of abdominal tumors
[101]. Another study showed that the delivery of miR-29a/c using cell-derived MVs inhibited angiogenesis in GC
[102]. Furthermore, EV-derived ncRNAs have been demonstrated to be functional towards tumor hallmarks in different cell lines. Huang et al. showed that exosomes derived from HCC cells silenced with circRNA-100338 could significantly decrease the invasive ability of HCC cells. In addition, these exosomes could reduce cell proliferation, angiogenesis, permeability, vasculogenic mimicry (VM) formation ability of HUVECs, and tumor metastasis
[83]. Bai et al. demonstrated that exosomal miR-135b secreted by GC cells inhibited the expression of FOXO1 protein and enhanced the growth of blood vessels in mouse models of tumor transplantation
[41]. Using an NPC model, Wang et al. found that overexpressed EBV-miR-BART10-5p and hsa-miR-18a upregulated VEGF and HIF-1α in a Spry3-dependent manner and strongly promoted angiogenesis. Moreover, in both in vitro and in vivo NPC models, treatment with iRGD-tagged exosomes enclosing antagomiR-BART10-5p and antagomiR-18a inhibited angiogenesis
[103]. Therefore, exosome engineering is a promising tool in RNA-based therapeutics for cancer treatment (
Figure 2B). Recently, RNA interference (RNAi)-based strategies, CRISPR/Cas9-mediated circRNA knockout, CRISPR/Cas13-mediated circRNA knockdown and circRNA-induced overexpressed plasmids were developed to target ncRNAs for therapeutic purposes both in vitro and in vivo
[104]. In a study, the expression of pro-angiogenic factors in HUVECs was significantly reduced after miR-92a-3p was knocked down in exosomes using an miR-92a-3p inhibitor (miR-92a-3p-i)
[105]. Furthermore, because EV-derived ncRNAs perform significant biological functions, specifically targeting EV-derived ncRNAs may be a promising strategy for treating many types of tumors. Currently, many studies aimed at regulating the production of EVs or blocking the uptake of EVs to achieve the goal of treating patients with cancer are underway. Using EVs as a delivery platform is a promising strategy; however, due to high costs and strict ethical regulations, the mass production of EVs is not easy to achieve to develop commercial viability.
This entry is adapted from the peer-reviewed paper 10.3390/cells11060947