The autonomic nervous system (ANS) and the immune system are deeply interrelated. The ANS regulates both innate and adaptive immunity through the sympathetic and parasympathetic branches, and an imbalance in this system can determine an altered inflammatory response as typically observed in chronic conditions such as systemic autoimmune diseases. Rheumatoid arthritis, systemic lupus erythematosus, and systemic sclerosis all show a dysfunction of the ANS that is mutually related to the increase in inflammation and cardiovascular risk. Moreover, an interaction between ANS and the gut microbiota has direct effects on inflammation homeostasis. Recently vagal stimulation techniques have emerged as an unprecedented possibility to reduce ANS dysfunction, especially in chronic diseases characterized by pain and a decreased quality of life as well as in chronic inflammation.
1. Introduction
The autonomic nervous system (ANS) has two main components, the sympathetic and the parasympathetic branches, that dynamically regulate the visceral functions [
1]. Previous and recent findings are confirming the strong reciprocal interrelation between ANS and the immune system. Since ANS can regulate inflammation in chronic and acute conditions, autonomic dysfunction can have a pivotal influence on the onset and progression of many diseases where the immune response is involved, such as autoimmune diseases [
2,
3,
4].
2. Autonomic Nervous System and Innate Immunity
The immune system is a complex interplay between immune cells, receptors, and self and non-self peptides. Innate immunity is the first response to microbes where pattern recognition receptors (PRRs) elicit the activation of immune/inflammatory processes after recognition of conserved pathogen-associated molecular patterns (PAMPs) that are present on bacteria, viruses, and fungi [
5] or towards damage- (or danger-) associated molecular patterns (DAMPs) and others. Among PRRs, Toll-like receptors (TLRs), nod-like receptors, C-type lectin receptors, and many others have a role in inducing innate immune responses; when PRRs are present on antigen-presenting cells, mainly dendritic cells (DCs), they can induce also the adaptive immune response [
6]. Furthermore, DCs play a pivotal role in contributing to either immune activation or maintaining the immune tolerance that is crucial to preventing autoimmunity [
7,
8]. Defensins, complement, granulocytes, and natural killer (NK) cells are components of innate immunity, determining inflammation that initially has a protective role from external or internal agents that have to be removed [
5,
9].
A deep interaction between the immune system and the nervous system is nowadays well documented and, in particular, innate immunity contributes to the development of the central nervous system (CNS) through microglia cells that are the main innate immune cells present in the brain [
10,
11]. TLRs are expressed on microglia surface responding to pathogenic or damaging insults [
12,
13,
14]. Furthermore, immune cells and innate immune cells maintain the functioning and homeostasis of the nervous system and an imbalance of this equilibrium, due for example to chronic inflammation, can cause a severe impairment with consequent alteration of cognitive functions [
15].
The interaction with innate immunity is not a prerogative of the CNS only and evidence show how the peripheral nervous system and more in specific the ANS have a deep interface with immune cells [
16]. Anatomically, the sympathetic branch of ANS is present in immunological organs such as the thymus, spleen bone marrow, and lymph nodes while of interest, no evident traces of the parasympathetic fibers have been demonstrated [
16,
17,
18,
19,
20,
21,
22]. Moreover, immune cells present adrenergic receptors able to bind norepinephrine that confers the ability to crosstalk with the sympathetic nerves, namely the alpha-adrenergic receptor (αAR) and the beta-adrenergic receptor (βAR), the latter more expressed on innate immune cells [
23,
24]. The receptors αAR and βAR have effects in opposite directions, where αAR can be considered more stimulatory, while βAR is inhibitory, and under homeostatic conditions, βAR has an overall predominant effect [
25]. Altogether, from several studies, norepinephrine inhibits cytokines production, namely TNFα when expressed from monocytes, macrophage, and microglia in response to the lipopolysaccharide (LPS) constituent of the bacterial cell-wall as well as inhibits IL-1β or IL-6 production [
26,
27,
28,
29,
30]. Norepinephrine has a direct effect on innate immune cells, increasing circulating NKs and granulocytes [
31,
32,
33]. Neutrophil chemotaxis and phagocytosis are negatively regulated from norepinephrine; the NK function impairment after stroke seems to be mediated by a noradrenergic neurotransmitter, and the NK response is suppressed by catecholamines [
34,
35,
36,
37,
38,
39,
40]. The βAR mediated effect of catecholamines suppress macrophage functions including their cytokine production [
41,
42]. Overall, the activation of the sympathetic nervous system attenuates the innate immunity as also demonstrated in a randomized control trial on human healthy subjects [
43].
The parasympathetic branch that includes the vagus nerve has several effects on the innate immune system through the interaction between receptors present on the cellular surface and neurotransmitters, namely acetylcholine [
44]. This communication is bidirectional and happens despite the fact that anatomically the parasympathetic fibers have not been individuated in the main immunologic organs such as the spleen and thymus [
16,
22]. Through vagal afferent fibers, the message that inflammation is present in other body sites reaches the CNS, as demonstrated by animal models of vagotomy in which the lack of vagal contribution determines reduced central responses with a blunted increase in body temperature and cortisol production [
45,
46,
47]. Evidence of how the vagal afferents are activated by inflammation is not yet completely clear, but it has been suggested that IL-1β receptors, present especially in the vagal paraganglia, are the main promotors of this afferent reflex to the CNS; moreover, IL-1β is itself a key contributor in the direct stimulation of the brain to activate the inflammatory cascade [
48,
49].
The vagus nerve has an anti-inflammatory effect through the release of acetylcholine, mainly through the interaction with the α7 nicotinic acetylcholine receptor (α7nAChR) present on macrophages [
50]. On cultures of LPS-stimulated human macrophages, acetylcholine attenuates the production of TNF, as well as of IL-6 and IL-1β, but not of the anti-inflammatory cytokine IL-10 [
51]. The spleen is one of the main targets of vagal action towards the immune system. Indeed, it has been demonstrated how vagal stimulation reduces TNF macrophage production in mice sepsis models [
52]. Due to the lack of parasympathetic fibers in the spleen, it has been hypothesized that the innervation is provided by catecholaminergic fibers from the celiac-superior mesenteric plexus ganglia that are under the control of preganglionic neurons of the thoracic spinal cord gray column [
19,
53,
54,
55]. Recently, an electrophysiological study performed on rats excluded the presence of a direct vagal-splenic nerve connection supporting the hypothesis of an effect towards splenic nerves mediated by vagal afferences through the CNS [
21,
33]. This neuronal modulation of inflammation through vagal afferences and efferences has been termed the “inflammatory reflex” [
56]. Overall, the inflammatory reflex is crucial to maintain homeostasis with a balance between pro and anti-inflammatory responses as evident by the increase in morbidity and mortality during sepsis when a vagal depression is present [
57,
58,
59,
60].
3. Autonomic Nervous System and Adaptive Immunity
Adaptive immunity is the specialized branch of immunity able to respond to specific pathogens and to maintain an immunological memory over time. The main cells involved are lymphocytes B and T. The sympathetic nervous system is able to regulate the mobilization of lymphocytes in the bloodstream through catecholamines that directly interact with β2AR present on the lymphocytes’ surface [
61]. Moreover, β2AR is selectively expressed on naïve T cells, CD4+ T helper (Th) 1, and regulatory T cells (Tregs) and induces T helper differentiation towards a Th1 phenotype through IFNγ/IL-12 interaction in in vitro studies, while in in vivo the Th differentiation is orchestrated via DCs+ [
62,
63]. Norepinephrine has an inhibitory effect on cytotoxic CD8+ T cells and modulates Tregs [
64,
65,
66]. Regarding B cells, catecholamines have an indirect effect on their maturation and on antibodies production through their action on T cells that are necessary as costimulation in the B mediated immune responses [
63]. Evidence on a direct effect of β2AR on B cells is limited; a lack of norepinephrine prevents a normal expression of IgG in mice [
67] and norepinephrine induces β2AR mediated CD86 expression (a costimulator) on B cells [
68,
69].
Vagal stimulation increases acetylcholine release in the spleen and suppresses TNF-α in control BALB/c mice models of endotoxemia, while it does not reduce TNF-α in nude mice, suggesting that T cells are involved in the inflammatory reflex and that a T cell deficiency impairs the inflammatory reflex [
70]. Moreover, α7nAChR present in T cells also causes a decrease in adhesion molecules expression and lymphocyte proliferation and both nicotinic and muscarinic acetylcholine receptors are present in lymphocytes that regulate their activities producing acetylcholine in a paracrine/autocrine control [
71,
72]. The role of vagal stimulation in increasing acetylcholine with beneficial effects on inflammation has been recently suggested also in the postural orthostatic tachycardia syndrome (POTS). POTS is a condition characterized by an impairment of the neuromodulation and consequent dysautonomia. Different studies showed a role of autoantibodies in POTS suggesting an autoimmune mediated pathogenesis of this condition [
73,
74]. In a recent study on a rabbit model of POTS induced by M2 muscarinic acetylcholine receptor-activating autoantibodies immunization, transcutaneous vagus nerve stimulation contributes to increasing acetylcholine with consequent reduction in both inflammation and cardiovagal dysfunction [
75].
Overall, once the inflammatory reflex is activated, the sympathetic and parasympathetic branches of ANS act synergistically instead of oppositely as intuitively expected
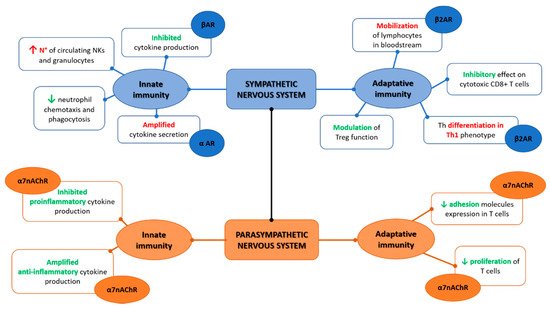
Figure 1. Indeed, as elegantly depicted by Tracey [
56], this synergic contribution implies that the vagal afferent fibers signal to the CNS (mainly within the nucleus of tractus solitarius) the presence of peripheric inflammatory/infective stimulus (intercepted for cytokines release and/or pathogens presence), and in response, vagal efferent fibers suppress cytokine release through nicotinic receptors present on macrophages, and throughout the cholinergic anti-inflammatory pathway. At the same time, the pain caused by the ongoing inflammatory processes can activate the sympathetic branches through the flight-or-fight responses determining norepinephrine release and consequent suppression of inflammation (via the pathways already detailed above) [
76,
77].
Figure 1. Sympathetic and parasympathetic synergic function on the innate and adaptive immunity.
It is important to add that these mechanisms can have different implications and functioning in acute versus chronic conditions, as described in acute stress that can cause an immune hyperactivation, while chronic stress is typically associated with an immunosuppressive status [
78], and what keeps the homeostasis is the dynamic balance between all these regulatory systems; when one system is prevailing, the imbalance can cause, or be the consequence of, a pathological condition, such as for example, chronic autoimmune diseases [
79]. Finally, by way of example of the deep bidirectional complex interactions between the nervous system and both innate and adaptive immunity, researchers could considered the case of celiac disease (CD) in which evidence shows how both innate and adaptive immunity mechanisms are involved [
80]. A wide range of neurological disorders, ANS dysfunction included, mediated by antineuronal and antigangliosides autoantibodies have been indeed demonstrated in CD [
81,
82,
83,
84].
4. Autonomic Nervous System and Gut Microbiota
The gastrointestinal tract (GIT) is considered one of the most extended and important immunological organs because of its enormous abundance of cells of both innate and adaptive immunity residing in the bowel mucosa [
85]. In the GIT, the immune system directly interacts with the unique microbiota ecosystems that are hosted there; microbiota includes the whole composition of bacteria, fungi, and viruses that are present in a specific body site, and the gut microbiota has a crucial role from birth, allowing the evolution and development of the immune system as demonstrated by germ-free mice models in which the absence of microbiota is associated with an absent or impaired immune development [
86,
87,
88,
89]. Moreover, GIT microbiota can regulate the immune interaction with external antigens and maintain the immune homeostasis through its protolerogenic commensal Phyla of bacteria able to metabolize and generate short-chain fatty acids (butyrate, propionate, and acetate) that induce Tregs expansion in the colon [
90,
91]. A reduction in pro-tolerogenic bacteria, mainly Firmicutes and Bacteroides has been extensively described in studies performed on mice models and in patients with inflammatory bowel diseases (IBD) and irritable bowel syndrome (IBS) as well as in systemic autoimmune diseases [
92,
93,
94,
95].
The brain–gut axis (BGA) is a well-known interaction between the enteric nervous system (ENS) and CNS that also occurs through the sympathetic and parasympathetic branches of ANS [
96,
97]. Gut microbiota can directly interact with the ENS and indirectly modulate the BGA through neuroendocrine and neuroimmune pathways, all together considered the “brain–gut–microbiota” axis [
98,
99]. If these mechanisms undergo a dysfunction, an imbalance of this system leads to clinical alteration of the GIT especially with IBS development [
100,
101,
102]. Moreover, the microbiota is directly associated with mental health disorders as demonstrated in knock-out mice models in which the absence of intestinal microbiota influences the development of behavior, along with neurochemical changes in the brain [
102,
103]. Microbiota alterations can modulate both the brain functions and the ANS through the vagus nerve, sending signals to the CNS and vice versa [
104,
105,
106,
107]. A recent study on beta 1 and 2 adrenergic receptor knock-out mice shows that the overall sympathetic reduction increases protolerogenic bacteria, with reduction in circulating CD4+ T cells and reduced IL-17 [
108].
This entry is adapted from the peer-reviewed paper 10.3390/ijms23052449