Anarhichotrema Shimazu, 1973 is a monotypic digenean genus, with the type- and only species, Anarhichotrema ochotense Shimazu, 1973, known to infect North Pacific fishes. This genus was originally described as a member of the Lissorchiidae (Monorchioidea) and later moved to the Zoogonidae (Microphalloidea).
1. Introduction
Anarhichotrema Shimazu, 1973 is a monotypic digenean genus erected by Shimazu [
1] for
A. ochotense Shimazu, 1973 described from the Bering wolffish
Anarhichas orientalis Pallas, 1814 (Anarhichadidae) caught in the Sea of Okhotsk. Twelve years later Machida [
2] described the genus
Neolissorchis Machida, 1985 with the type and only species
Neolissorchis genge Machida, 1985 from the intestine of Zoarcid fish
Lycodes sp. in the Sea of Japan (East Sea). Machida [
2] was apparently unaware of the publication by Shimazu [
1] since he did not mention
A. ochotense in his work. However, the two species had extremely similar morphology. Thereafter, Bray [
3] reasonably synonymised
Neolissorchis with
Anarhichotrema and
N. genge with
A. ochotense. Later Kuramochi [
4] found
A. ochotense in a Zoarcid fish
Petroschmidtia toyamensis Katayama, 1941 (=
Lycodes toyamensis (Katayama, 1941)) in the Sea of Japan (East Sea).
Since
Anarhichotrema is similar to
Lissorchis Magath, 1917, the type-genus of the Lissorchiidae Magath, 1917, in general morphology, it was originally described as a member of the Lissorchiidae [
1], and so was
Neolissorchis [
2]. However, Bray [
3,
5] moved
Anarhichotrema into the Zoogonidae based on the understanding that lissorchiids did not typically parasitise marine fish. According to the current zoogonid concept,
Anarhichotrema is affiliated to the Lecithostaphylinae Odhner, 1911
sensu lato [
6]. However, the lack of molecular genetic data for this genus significantly complicates evaluation of its phylogenetic position.
2. Phylogenetic Relationships
The 28S rRNA gene sequences were almost identic to each other after trimming and alignment. Only one gap was observed in position 1266 of general alignment in sequence OM108706. The BLAST hit results showed that the 28S rRNA sequences of A. ochotense were more similar to those of lissorchiid and monorchiid digeneans.
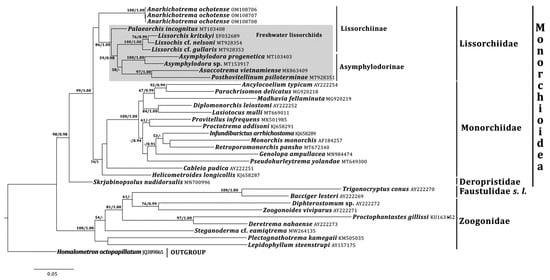
Maximum likelihood and Bayesian Inference analyses based on 28S rRNA gene data placed A. ochotense within the Monorchioidea, sister to the well-supported ‘freshwater lissorchiids’ clade; the position of A. ochotense was poorly-supported (Figure 1). The ‘freshwater lissorchiids’ clade included the paraphyletic assemblage of the lissorchiine digeneans and the monophyletic Asymphylodorinae. However, the Asymphylodorinae clade was not well-supported in any of the analyses. In turn, the A. ochotense + ‘freshwater lissorchiids’ clade resolved sister to the Monorchiidae; this larger clade was well-supported. The degree of divergence between the 28S rRNA gene sequences of A. ochotense and the basal species in the ‘freshwater lissorchiids’ clade (Palaeorchis incognitus Szidat, 1943) did not exceed that between the basal monorchiids (Cableia pudica Bray, Cribb & Barker, 1996, Helicometroides longicollis Yamaguti, 1934), being 10% and 17%, respectively. In the trees based on 18S+28S rRNA gene data, A. ochotense formed a well-supported clade with Lissorchis kritskyi Barnhart & Powell, 1979 (Figure 2).
Figure 1. Phylogenetic relationships of Anarhichotrema ochotense based on dataset of 28S rRNA gene sequences. The bootstrap (≥50) and posterior probability (≥0.90) values are given near the nodes for ML and BI analyses, respectively.
Figure 2. Phylogenetic relationships of Anarhichotrema ochotense based on concatenated dataset of 18S+28S rRNA gene sequences. The bootstrap (≥50) and posterior probability (≥0.90) values are given near the nodes for ML and BI analyses, respectively.
3. Systematics
Anarhichotrema ochotense Shimazu, 1973 Figure 3
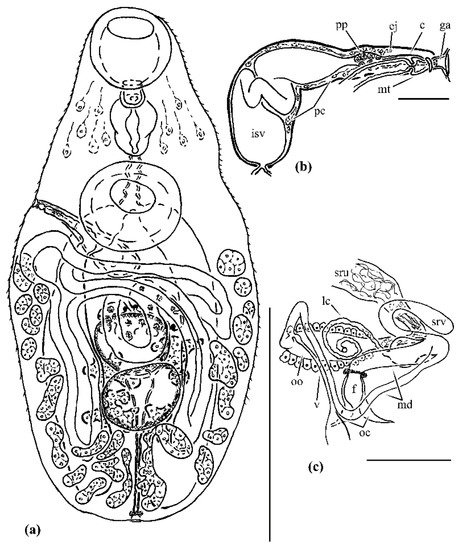
Figure 3. Anarhichotrema ochotense from intestine of Anarhichas orientalis, the Sea of Okhotsk: (a) whole mount, ventral view; (b), terminal genitalia, ventral view; (c), ovarian complex, dorsal view. Abbreviations: c, invaginated cirrus; ej, ejaculatory duct; f, fertilization chamber; ga, common genital atrium; isv, internal seminal vesicle; lc, Laurer’s canal; md, middle part of oviduct; mt, metraterm; oc, ovicapt; oo, oötype with Mehlis’ gland cells; pc, prostatic cells; pp, pars prostatica; src, canalicular seminal receptacle; sru, uterine seminal receptacle; v, vitelline reservoir. Scale bars: (a) = 500 µm, (b,c) = 50 µm.
Body fusiform, maximum width near mid-level of body. Tegument spined. Forebody with numerous gland-cells in parenchyma. Oral sucker round, mouth subterminal. Ventral sucker rounded, unspecialised. Prepharynx short, muscular. Pharynx muscular, elongate-oval. Oesophagus contracted to some extent in all specimens. Intestinal bifurcation dorsal to ventral sucker. Caeca blind, terminate near posterior extremity of body.
Testes two, irregularly indented (in four specimens) or entire (in one specimen), tandem, slightly overlapping, in posterior half of body. Cirrus sac rectilinear to comma-shaped, passes beyond posterior margin of ventral sucker, terminating in a common genital atrium. Vas deferens absent; vasa efferentia directly joining seminal vesicle. Internal seminal vesicle unipartite, tubular, sinuous. Pars prostatica vesicular; lined with anuclear cell-like bodies; field of prostatic cells extensive. Ejaculatory duct rectilinear, invaginated cirrus unarmed. Common genital atrium small, distinct. Genital pore sinistro- (in two specimens) or dextro-marginal (in three specimens), located near midlevel of ventral sucker.
Ovary multilobate, median, overlapping anterior edge of anterior testis. Ovicapt distinct. Proximal part of oviduct forms thick-walled fertilization chamber terminating with sphincter. Middle part of oviduct long, linked to canalicular seminal receptacle and Laurer’s canal, and vitelline reservoir. Distal part of oviduct forms oötype. Canalicular seminal receptacle slender, without sperm. Laurer’s canal bulbous proximally, opens medially, anterior to ovary. Oötype with compact Mehlis’ gland, antero-ventral to ovary. Uterus tortuous, reaching into post-testicular region, proximal portion functions as uterine seminal receptacle. Metraterm short, separated from uterus by constriction, terminating with sphincter, unarmed, ventral to cirrus sac. Eggs numerous, operculate, with long anopercular filament. Vitellarium follicular; follicles form two ventrolateral fields and median dorsal group. Ventrolateral fields extend from level of posterior edge or posterior third of ventral sucker to posterior extremity of body, confluent in post-testicular region. Follicles of dorsal group in two submedian longitudinal rows; rows asymmetrical in most specimens, anterior-most follicle at midlevel of anterior testis to midpoint of ventral sucker-ovary distance, posterior-most follicle at level of posterior edge of posterior testis or anterior third or middle of post-testicular region or near posterior extremity of body. Vitelline reservoir median, ventral to ovary.
Excretory vesicle I-shaped, extends to middle or anterior edge of posterior testis, posterior end surrounded by small muscular sphincter; excretory pore terminal.
4. The Results of Phylogenetic Analysis
The specimens of A. ochotense described are similar to those from Shimazu [1] in most of the morphological features related to the shape and location of the organs. Only three features appear to be different, based on the drawing in Shimazu [1]: the relative position of the testes (overlapping vs. separate), the position of the anterior border of the ventrolateral fields of the vitelline follicles (at the level of or slightly anteriorly to the posterior edge of the ventral sucker vs. distinctly posterior to the ventral sucker), and the arrangement of the cirrus sac (overlapped by the ventral sucker vs. lying outside the projection of the sucker). However, Machida [2] demonstrated that these features can be variable.
At the same time, the trematode specimens were noticeably smaller than those studied by T. Shimazu and M. Machida. This difference concerned all morphometric characteristics except egg length. The small size of the parasites could be associated with the crowding effect: A. ochotense infection intensity was 375 individuals as compared with six individuals in Shimazu [1] and five individuals in Machida [2]. The so-called crowding effect is a result of competition for host‘s finite resources by parasites. Typically, this effect increases with increasing numbers of parasites within a host and manifests in reduced body size and thus fitness [37,38,39].
Phylogenetic analyses indicate that Anarhichotrema is a member of the Lissorchiidae [1].The family Lissorchiidae is currently divided into two subfamilies, the Lissorchiinae Magath, 1917 and the Asymphylodorinae Szidat, 1943 [5]. The division is based on the number of testes: two in the former and one in the latter subfamily [5]. Phylogenetic data support the unification of the lissorchiine and the asymphylodorine digeneans into the same family but do not support the monophyly of the Lissorchiinae ([9,10,11,40], Present Study). Notably, the position of two lissorchiine genera, Lissorchis and Palaeorchis Szidat, 1943, is unstable in the phylograms of different authors [10,11], Present Study. Anarhichotrema, in contrast to the other lissorchiid genera, has a marine life history and the filamented eggs compare with [5,9,11]. Taking into account the ecological and morphological features of Anarhichotrema as well as the phylogenetic data (the basal position of Anarhichotrema and the paraphyly of the Lissorchiinae), it is conjectured that a separate subfamily may have to be erected for this genus in the future and that all freshwater lissorchiids should probably be considered within one subfamily (Lissorchiinae). However, for now, Anarhichotrema to the Lissorchiinae is conditionally considered.
This entry is adapted from the peer-reviewed paper 10.3390/d14020147