The constitutively active BCR-ABL1 tyrosine kinase, found in t(9;22)(q34;q11) chromosomal translocation-derived leukemia, initiates an extremely complex signaling transduction cascade that induces a strong state of resistance to chemotherapy. Targeted therapies based on tyrosine kinase inhibitors (TKIs), such as imatinib, dasatinib, nilotinib, bosutinib, and ponatinib, have revolutionized the treatment of BCR-ABL1-driven leukemia, particularly chronic myeloid leukemia (CML). However, TKIs do not cure CML patients, as some develop TKI resistance and the majority relapse upon withdrawal from treatment. Importantly, although BCR-ABL1 tyrosine kinase is necessary to initiate and establish the malignant phenotype of Ph-related leukemia, in the later advanced phase of the disease, BCR-ABL1-independent mechanisms are also in place.
1. Introduction
BCR-ABL1 is a multidomain, constitutively active, chimeric tyrosine kinase that results from a reciprocal translocation between chromosomes 9 and 22—t(9;22)(q34;q11)—characteristic of Philadelphia chromosome(Ph1)-positive leukemia [
1]. Depending on the breakpoint on chromosome 22 at the
BCR (break point cluster) gene, three major isoforms of BCR-ABL1 can be produced: the 185kDa, 210kDa, and 230kDa proteins found in acute lymphocytic leukemia (ALL), chronic myeloid leukemia (CML), and chronic neutrophilic leukemia (CNL), respectively [
2,
3,
4]. In all these circumstances, the first exon of
c-ABL—the cellular homolog of Abelson murine leukemia virus (A-MuLV)—on chromosome 9 is replaced by one of the
BCR sequences (
Figure 1).
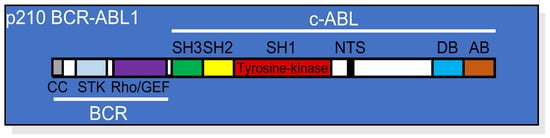
Figure 1. Linear structure of p210 BCR-ABL1 showing the relative position of each domain from both the BCR and the c-ABL portions of the protein. CC, coil-coiled oligomerization domain; STK, serine/threonine kinase domain; Rho/GEF domain; SH, (SRC homology domains) 1, 2, and 3; NTS, nuclear translocation signal; DB, DNA binding domain; AB, actin binding domain.
The BCR part of the protein contributes to several domains responsible for regulating the enzymatic activity of BCR-ABL1 or its interactions with different binding partners [
5,
6]. At the N-terminal portion of BCR, there is a coiled-coil domain responsible for oligomerization and constitutive activation of the BCR-ABL1 tyrosine kinase. In addition, the BCR sequence contains a serine/threonine kinase (STK) domain, a Ras homolog gene family/guanine nucleotide exchange factors (Rho/GEF) kinase domain, and SH2 domains capable of binding adaptor molecules, such as growth factor receptor-bound protein 2 (GRB2) [
7].
c-ABL is a tyrosine kinase located preferentially in the nucleus [
8], although it is also found in the cytoplasm where it associates with actin filaments [
5]. The structure of c-ABL, which is conserved in BCR-ABL1, comprises multiple domains, including SRC-homology domain 1 (SH1; kinase domain), SH2, SH3, DNA binding (DB), and actin binding (AB) domains, in addition to a nuclear translocation signal (NTS) sequence, sites for phosphorylation by protein kinase C (PKC), and a proline-rich sequence (
Figure 1). Among all these domains, the SH1 region is the most conserved during evolution and contains the catalytic site essential for the initiation of signaling pathways that result in cellular transformation, including dysregulated proliferation and resistance to apoptosis.
2. BCR-ABL1 Tyrosine Kinase-Dependent Signaling Cascade
Due to its complex structural nature, multiple proteins have been shown to directly associate with BCR-ABL1. Co-immunoprecipitation of BCR-ABL1 from the K562 CML cell line followed by mass spectrometry revealed the potential core components of the BCR-ABL1-interactome, including the adapter proteins GRB2, SHC adaptor protein 1 (SHC1), CT10 regulator of kinase 1 (CRK1), the E3 ubiquitin-protein ligase casitas B-lineage lymphoma (c-CBL), the p85α and β subunits of the phosphoinositide 3-kinase (PI3K), the suppressor of T-cell receptor signaling 1 (STS1), and the SH2 domain-containing inositol 5-phosphatase 2 (SHIP-2) [
14]. These proteins, along with other interactors, can recruit intermediate and/or effector molecules, thereby initiating a plethora of signaling pathways, including RAS/RAF/MAPK, PI3K/AKT/mTOR, JAK/STAT, and WNT/β-catenin, responsible for the different aspects of BCR-ABL-1-induced transformation.
2.1. BCR-ABL1 Activation of the RAS/RAF/MAPK Pathway
The activation of the rat sarcoma virus (RAS)/rapidly accelerated fibrosarcoma (RAF)/mitogen-activated protein kinase (MAPK) pathway by BCR-ABL1 is initiated by GRB2 binding to phosphotyrosine Y177 in the BCR region, followed by the recruitment of GRB2-associated-binding protein 2 (GAB-2) and son of sevenless (SOS). The BCR–ABL1–GRB2–SOS complex drives RAS activation and the consequent activation of RAF1, MAPK/ERK Kinase (MEK), extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38MAPK. Together, these signaling pathways regulate cell proliferation, differentiation, and survival [
15,
16,
17,
18].
2.2. BCR-ABL1 Activation of the PI3K/AKT/mTOR Pathway
BCR-ABL1 activates the PI3K/AKT/mTOR pathway both directly and indirectly through the induction of autocrine cytokines [
21]. The interaction between BCR-ABL1 and PI3K can occur via GRB2, GAB-2, SHC, c-CBL, and CRKL [
15,
22,
23].
AKT also induces the downstream activation of mammalian target of rapamycin (mTOR), which works as a catalytic subunit of the mTORC1 and mTORC2 protein complexes [
31].
2.3. BCR-ABL1 Activation of the JAK/STAT Pathway
Janus kinase (JAK)/signal transducer and activator of transcription (STAT) signaling is involved in growth factor independence and resistance to apoptosis in CML [
43,
44,
45,
46,
47]. JAK2, STAT1, STAT3, and STAT5 were shown to be constitutively active in CML cell lines [
47].
2.4. BCR-ABL1 Activation of the WNT/β-Catenin Pathway
A disturbance in the canonical WNT/β-catenin signaling pathway is associated with the pathogenesis of leukemia [
53,
54]. In the absence of WNT activation, β-catenin is phosphorylated and ubiquitinated in the context of a multiprotein complex composed of glycogen synthase kinase 3 (GSK-3), creatin kinase 1 (CK1), axis inhibitor (Axin), adenomatous polyposis coli (APC), protein phosphatase 2A (PP2A), and the E3-ubiquitin ligase β-transducin repeat-containing protein (β-TrCP), followed by further proteasomal degradation [
53,
54].
2.5. BCR-ABL1 Activation of the PP2A Pathway
PP2A is a tumor suppressor serine-threonine phosphatase that negatively regulates the mitogenic and survival signals emanating from PI3K/AKT, RAS/MAPK, and MYC pathways [
59].
3. BCR-ABL1 Kinase-Independent Alternative Survival Signals
Unquestionably, the BCR-ABL1 tyrosine kinase-dependent signaling events are required for the transformation of Ph1 chromosome-positive leukemia. However, the resistance to TKIs observed in some CML patients suggests that signals emanating from the BCR-ABL1 protein independently of its tyrosine kinase activity may take over, allowing the survival of leukemic cells and relapse of the disease. Indeed, it was recently shown that inhibition of the catalytic activity does not completely dismantle the BCR-ABL1 molecular complex [
63]. Signaling proteins, such as p85α-PI3K, GRB2, SHIP2, SHC1, SOS1, and c-CBL, remain associated with BCR-ABL1, whereas CRK, CRKL, or GAB2 seem to detach from the complex [
63]. Therefore, residual signaling transduction events appear to be sufficient to maintain survival of CML cells in the absence of tyrosine kinase activity [
64].
4. Resistance to Apoptosis in CML
BCR-ABL1 activates multiple signaling pathways to induce leukemogenesis, which results in growth factor-independency and regulation of adhesion and invasion [
71,
72,
73]. On the other hand, BCR-ABL1-positive cells generally display normal mitotic indices and do not show increased overall proliferation [
74]. Perhaps the most notable aspect of BCR-ABL1-mediated leukemogenesis is the vigorous state of resistance to apoptosis that is conferred to the transformed cells [
19,
75,
76,
77,
78,
79,
80].
The enforced expression of BCR-ABL1 in hematopoietic cell lineages revealed its potential to prevent apoptosis induced by a variety of stimuli, including growth factor withdrawal, γ-irradiation, death receptor agonists, and multiple chemotherapeutic drugs [
19,
75,
76,
77,
78,
79]. Studies with point mutations at the autophosphorylation site (Y793F), the phosphotyrosine binding motif (R552L), and/or at the GRB2-binding site (Y177F) demonstrated that BCR-ABL1-mediated resistance to apoptosis depends on the cellular context. For instance, enforced expression of a BCR-ABL1-Y177F/R552L/Y793F triple mutant in IL-3-dependent lymphoblastoid 32D murine cells did not confer IL-3 independency or resistance to γ-irradiation-induced apoptosis; however, the same mutant protected BaF3 cells, a different IL-3-dependent pro-B murine cell line, from these apoptogenic stimuli [
19]. Moreover, enforced expression of wild type or the above-mentioned BCR-ABL1 mutants protected the apoptosis-sensitive human acute promyelocytic leukemia HL-60 cell line from a variety of apoptogenic insults to the same extent [
77]. Therefore, different cellular contexts may provide alternative pathways that contribute to the survival of BCR-ABL1-positive cells.
5. Tyrosine Kinase Inhibitors and the Paradigm Shift of CML Treatment
The discovery that constitutive BCR-ABL1 tyrosine kinase activity was crucial for the development of CML [
96] warranted a TK-targeting therapeutic strategy. Consequently, several TKIs were developed to target the ATP binding site of the kinase domain, thereby preventing phosphorylation of the target protein and subsequent signaling events (
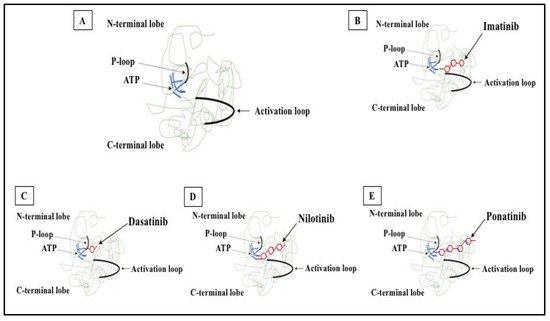
Figure 2). Imatinib, nilotinib, dasatinib, bosutinib, ponatinib, and asciminib are currently used for the treatment of CML and are briefly described below.
Figure 2. 2D structure of BCR-ABL1 kinase domain and binding sites for TKIs. (A) BCR-ABL1 is a constitutively active kinase that binds ATP and transfers a phosphate from ATP to tyrosine residues on various substrates. This activates downstream signaling pathways, leading to abnormal cellular adhesion and proliferation of myeloid cells and inhibition of apoptosis. TKIs were developed to specifically block the binding of ATP to the BCR-ABL tyrosine kinase, inactivating the constitutive tyrosine kinase activity and inhibiting downstream pathways. (B) Imatinib (first generation TKI) binds to the BCR-ABL kinase domain in its inactive conformation through the ATP binding site. (C) Dasatinib (second generation TKI) inhibits the BCR-ABL tyrosine kinase performance at the ATP site in ABL regardless of protein conformation (active or inactive). (D) Nilotinib (second generation TKI) connects to an inactive conformation of the BCR-ABL protein, taking an analogous region that would be occupied by ATP. (E) Ponatinib (third generation TKI) has multiple contact points for the inactive conformation of the ABL and for the T315I mutation.
6. Mechanisms of Resistance to TKI
The major purpose of CML therapy with TKIs is to achieve a deep molecular response. To identify those who have not achieved desired response and may be suffering from resistance to TKIs, it is necessary to apply the European LeukemiaNet criteria for treatment response [
124,
127]. To maintain the current treatment, an optimal response is mandatory; otherwise, the therapy must be replaced (failure/resistance) or considered for change (warning) [
124]. Furthermore, to achieve successful treatment, it is necessary to understand the mechanisms that led to the poor outcome, particularly the resistance to TKIs (
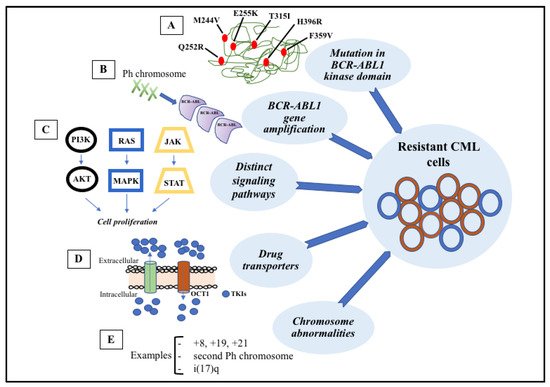
Figure 3).
Figure 3. Signaling pathways involved in the development of target therapy resistance. (A) Molecular structure of BCR-ABL1 kinase domain with some mutations (indicated in red). (B) Gene amplification can lead to overproduction of tyrosine kinase. (C) Constitutive activation of signaling pathways, such as PI3K-AKT, RAS-MAPK, and JAK-STAT, result in cell proliferation and anti-apoptotic mechanisms. (D) Intracellular concentrations of TKIs can be modified through membrane transporters that may cause increased efflux or decreased influx. (E) The most common chromosome abnormalities involved in karyotype evolution are trisomy 8, trisomy 19, trisomy 21, second Ph chromosome, and isochromosome 17.
Resistance to TKIs can be classified as primary (no hematologic or cytogenetic response from the beginning of therapy) or secondary (initial response that decays during the treatment). Currently, two mechanisms of resistance are known: BCR-ABL1-dependent and BCR-ABL1-independent pathways [
122]. It is important to note that the ATP molecule binds between the two lobes of the catalytic domain in ABL kinase (an N-terminal lobe and a C-terminal lobe) [
128]. Mutations that lead to imatinib resistance have been detected in the phosphate-binding loop and in other regions of the kinase domain where amino acid substitutions may result in conformational changes that prevent imatinib binding [
129].
This entry is adapted from the peer-reviewed paper 10.3390/pharmaceutics14010215