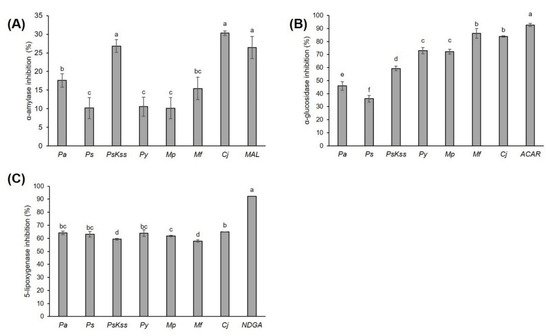
This work aims to assess the biopotential of the young inflorescence tissues of Prunus, Malus and Chaenomeles in order to evaluate the possibility of their application in the food industry, and to provide a polyphenolic fingerprint for their quality control. The contents of different bioactive compounds and their antioxidant capacities were spectrophotometrically measured, the main phenolic compounds were identified and quantified using LC-DAD-MS, the antidiabetic potential was determined using α-amylase and α-glucosidase inhibition assays, the anti-inflammatory potential was determined using a 5-lipoxygenase inhibition assay, and the cytotoxicity was determined by MTT assay. Using one-way ANOVA, principal component analysis, hierarchical clustering and Pearson’s correlation coefficient, the relations between the samples, and between the samples and the measured parameters, were revealed. In total, 77 compounds were identified. The concentration of sugars was low in M. purpurea, at 1.56 ± 0.08 mg/g DW. The most effective sample in the inhibition of antidiabetic enzymes and anti-inflammatory 5-lipoxygenase was C. japonica. The inhibition of α-glucosidase was strongly positively correlated with the total and condensed tannins, procyanidin dimers and procyanidin tetramer, and was very strongly correlated with chlorogenic acid. In α-amylase inhibition, C. japonica and P. serrulata ‘Kiku Shidare Zakura’ were equally efficient to the standard inhibitor, maltose. The most effective in the growth and proliferation inhibition of HepG2, HCT116 and HaCaT cells was P. avium. The results suggest Prunus, Malus and Chaenomeles inflorescences as functional food ingredients.
- 5-lipoxygenase
- α-amylase
- α-glucosidase
- Chaenomeles
- colorectal carcinoma
- hepatocellular carcinoma
- keratinocytes
- Malus
- metabolomics
- Prunus
1. Introduction
2. Spectrophotometric Analysis of the Phytochemical Content and Antioxidant Capacity of the Inflorescences
| Prunus avium | Prunus serrulata | Prunus serrulata ‘Kiku Shidare Zakura’ | Prunus yedoensis | Malus purpurea | Malus floribunda | Chaenomeles japonica | |
|---|---|---|---|---|---|---|---|
| TP (mg GAE/g DW) | 27.83 ± 0.69 f | 46.84 ± 0.75 b | 53.12 ± 0.79 a | 37.31 ± 0.48 e | 40.29 ± 0.64 c | 46.74 ± 0.93 b | 39.42 ± 0.97 d |
| TF (mg CE/g DW) | 13.57 ± 0.84 f | 32.35 ± 2.13 b | 38.89 ± 4.04 a | 25.78 ± 1.03 d | 13.43 ± 0.82 f | 23.83 ± 0.88 e | 29.45 ± 0.65 c |
| TNF (mg GAE/g DW) | 16.40 ± 0.91 g | 29.35 ± 0.83 b | 31.32 ± 0.71 a | 22.21 ± 0.86 e | 23.52 ± 0.56 d | 28.85 ± 0.58 c | 18.73 ± 0.48 f |
| TT (mg CE/g DW) | 27.26 ± 0.22 g | 59.44 ± 1.32 f | 71.59 ± 0.33 d | 83.55 ± 0.55 b | 64.32 ± 0.45 e | 107.85 ± 1.09 a | 80.27 ± 0.33 c |
| CT (mg CE/g DW) | 4.25 ± 0.33 e | 7.74 ± 1.02 d | 6.99 ± 0.17 d | 16.52 ± 0.01 b | 15.45 ± 0.10 b | 10.98 ± 0.24 c | 51.68 ± 0.38 a |
| SS (mg SE/g DW) | 3.37 ± 0.06 c | 3.04 ± 0.05 d | 2.41 ± 0.10 e | 8.61 ± 0.12 a | 1.56 ± 0.08 f | 3.58 ± 0.06 b | 3.27 ± 0.07 c |
| ABTS (mg TE/g DW) | 22.86 ± 4.61 e | 49.41 ± 7.23 b | 61.32 ± 5.84 a | 36.63 ± 4.32 c | 28.78 ± 2.52 d | 47.78 ± 6.26 b | 35.05 ± 4.25 c |
| FRAP (mg TE/g DW) | 27.89 ± 0.60 g | 51.68 ± 0.12 b | 58.06 ± 0.78 a | 40.28 ± 1.18 d | 29.12 ± 0.83 f | 44.36 ± 0.85 c | 36.36 ± 1.50 e |
| DPPH (mg TE/g DW) | 25.47 ± 2.57 d | 52.95 ± 4.22 b | 69.42 ± 3.27 a | 39.21 ± 4.86 c | 25.10 ± 3.25 d | 40.38 ± 3.4 c | 39.61 ± 3.95 c |
| Prunus avium | Prunus serrulata | Prunus serrulata ‘Kiku Shidare Zakura’ | Prunus yedoensis | Malus purpurea | Malus floribunda | Chaenomeles japonica | ||
|---|---|---|---|---|---|---|---|---|
| 1 | Gallic acid | 0.28 ± 0.06 b | 0.15 ± 0.04 c | 0.06 ± 0.01 d | 0.38 ± 0.03 a | 0.17 ± 0.04 c | 0.03 ± 0.01 d | nd |
| Total identified hydroxybenzoic acids |
0.28 ± 0.06 b | 0.15 ± 0.04 c | 0.06 ± 0.01 d | 0.38 ± 0.03 a | 0.17 ± 0.04 c | 0.03 ± 0.01 d | nd | |
| 2 | Caffeic acid | 1.95 ± 0.27 c | 1.25 ± 0.14 c | 2.50 ± 0.08 b | 3.90 ± 0.10 a | nd | nd | 0.53 ± 0.08 d |
| 3 | Caffeic acid hexoside 1 | 0.14 ± 0.03 b | 0.39 ± 0.06 b | 15.48 ± 2.23 a | 0.10 ± 0.01 b | 0.05 ± 0.01 b | 0.23 ± 0.02 b | 0.75 ± 0.04 b |
| 4 | Caffeic acid hexoside 2 | nd | nd | nd | 5.62 ± 0.25 a | nd | nd | 0.02 ± 0.00 b |
| 5 | Caffeic acid dihexoside | 0.29 ± 0.01 b | 0.19 ± 0.04 c | 0.41 ± 0.02 a | nd | nd | nd | nd |
| 6 | 3-caffeoylquinic acid | nd | 0.36 ± 0.04 b | 1.70 ± 0.25 a | 0.54 ± 0.03 b | nd | nd | 0.02 ± 0.00 c |
| 7 | 4-caffeoylquinic acid | nd | 0.27 ± 0.08 a | 0.35 ± 0.02 a | nd | 0.13 ± 0.06 b | 0.15 ± 0.02 b | 0.07 ± 0.01 b |
| 8 | 5-caffeoylquinic acid 1 | 1.59 ± 0.18 d | 0.59 ± 0.17 e | 0.47 ± 0.02 e | 5.75 ± 0.15 c | 1.41 ± 0.30 d | 8.41 ± 0.69 a | 7.04 ± 0.23 b |
| 9 | 5-caffeoylquinic acid 2 | 0.43 ± 0.36 a | nd | nd | 0.27 ± 0.01 a | 0.31 ± 0.13 a | nd | 0.14 ± 0.02 a |
| 10 | di-caffeoylquinic acid 1 | 3.12 ± 0.08 b | 0.15 ± 0.03 c | 0.31 ± 0.02 c | 7.06 ± 0.82 a | 0.65 ± 0.14 c | 0.50 ± 0.03 c | 2.83 ± 0.30 b |
| 11 | di-caffeoylquinic acid 2 | 0.13 ± 0.01 b | nd | nd | 0.29 ± 0.01 b | 0.16 ± 0.06 b | 1.02 ± 0.15 a | nd |
| 12 | di-caffeoylquinic acid 3 | 0.17 ± 0.03 a | nd | nd | nd | nd | nd | nd |
| 13 | 3-feruloylquinic acid | 0.24 ± 0.04 a | 0.03 ± 0.00 c | 0.05 ± 0.00 bc | 0.08 ± 0.00 b | nd | nd | 0.004 ± 0.001 c |
| 14 | 5-feruloylquinic acid | 0.26 ± 0.03 a | 0.06 ± 0.01 d | 0.11 ± 0.00 bc | 0.09 ± 0.01 cd | 0.01 ± 0.00 e | 0.13 ± 0.04 b | 0.01 ± 0.00 e |
| 15 | 3-p-coumaroylquinic acid | 0.52 ± 0.08 b | 0.14 ± 0.02 d | 0.21 ± 0.01 c | 0.73 ± 0.03 a | 0.03 ± 0.01 e | 0.15 ± 0.01 cd | 0.001 ± 0.000 e |
| 16 | 4-p-coumaroylquinic acid | nd | 0.27 ± 0.08 a | 0.35 ± 0.02 a | nd | 0.13 ± 0.06 b | 0.15 ± 0.02 b | 0.07 ± 0.02 b |
| 17 | 5-p-coumaroylquinic acid 1 | 0.39 ± 0.02 a | 0.09 ± 0.01 d | 0.07 ± 0.00 d | 0.33 ± 0.03 b | 0.10 ± 0.01 d | 0.19 ± 0.03 c | 0.12 ± 0.03 d |
| 18 | 5-p-coumaroylquinic acid 2 | 0.03 ± 0.00 c | 0.09 ± 0.01 b | 0.08 ± 0.01 bc | 0.10 ± 0.01 b | 0.07 ± 0.06 bc | 0.06 ± 0.02 bc | 0.25 ± 0.04 a |
| 19 | p-coumaric acid hexoside 1 | 0.23 ± 0.06 b | 0.21 ± 0.04 b | 0.31 ± 0.01 a | 0.09 ± 0.00 c | 0.04 ± 0.01 cd | 0.22 ± 0.04 b | 0.001 ± 0.000 d |
| 20 | p-coumaric acid hexoside 2 | 0.37 ± 0.06 b | 1.42 ± 0.15 a | 1.34 ± 0.05 a | 0.21 ± 0.01 c | 0.02 ± 0.00 d | 0.11 ± 0.01 cd | 0.13 ± 0.00 cd |
| Total identified hydroxycinnamic acids |
9.87 ± 0.08 b | 5.51 ± 0.06 c | 23.70 ± 0.18 a | 25.16 ± 0.10 a | 3.12 ± 0.06 d | 11.31 ± 0.09 b | 11.93 ± 0.05 b | |
| 21 | Catechin | 0.23 ± 0.04 e | 2.61 ± 0.27 a | 2.46 ± 0.09 a | 0.91 ± 0.02 d | nd | 1.99 ± 0.16 b | 1.67 ± 0.06 c |
| 22 | Epicatechin | 1.10 ± 0.15 d | 0.91 ± 0.10 d | 1.81 ± 0.06 b | 6.92 ± 0.18 a | 1.18 ± 0.18 d | 1.38 ± 0.22 c | 0.40 ± 0.06 e |
| Total identified flavanols | 1.33 ± 0.09 e | 3.52 ± 0.18 cd | 4.27 ± 0.07 b | 7.83 ± 0.10 a | 1.18 ± 0.18 e | 3.38 ± 0.19 bc | 2.08 ± 0.06 de | |
| 23 | Eriodictyol hexoside 1 | 0.03 ± 0.00 b | nd | nd | nd | 0.63 ± 0.04 b | 2.92 ± 0.86 a | nd |
| 24 | Eriodictyol hexoside 2 | nd | nd | nd | nd | 0.35 ± 0.09 b | 1.02 ± 0.15 a | nd |
| 25 | Naringenin hexoside | nd | nd | nd | nd | nd | nd | 0.62 ± 0.04 a |
| Total identified flavanones | 0.03 ± 0.00 b | nd | nd | nd | 0.98 ± 0.06 b | 3.93 ± 0.50 a | 0.62 ± 0.04 b | |
| 26 | Quercetin-glycoside | 0.18 ± 0.01 c | 0.13 ± 0.01 d | 0.21 ± 0.02 b | 0.36 ± 0.02 a | nd | nd | nd |
| 27 | Quercetin-3-rutinoside | 2.78 ± 0.28 b | 0.58 ± 0.10 d | 4.79 ± 0.13 a | 1.93 ± 0.03 c | 0.08 ± 0.03 e | 0.21 ± 0.03 e | 0.72 ± 0.04 d |
| 28 | Quercetin-3-rhamnoside hexoside | nd | nd | nd | nd | 0.58 ± 0.21 a | 0.41 ± 0.06 a | nd |
| 29 | Quercetin-hexoside pentoside | nd | 0.25 ± 0.08 b | 0.59 ± 0.01 a | nd | nd | nd | nd |
| 30 | Quercetin-rhamnoside dihexoside 1 | 0.15 ± 0.01 b | nd | nd | 0.20 ± 0.00 a | nd | nd | nd |
| 31 | Quercetin-rhamnoside dihexoside 2 | nd | 0.02 ± 0.01 c | 0.05 ± 0.00 b | 0.47 ± 0.01 a | nd | nd | nd |
| 32 | Quercetin-3-galactoside | 0.26 ± 0.02 c | nd | nd | 0.98 ± 0.01 b | 1.21 ± 0.27 a | 0.309 ± 0.070 c | 0.17 ± 0.01 c |
| 33 | Quercetin-3-glucoside | 0.03 ± 0.00 d | 0.19 ± 0.02 a | 0.12 ± 0.01 b | nd | 0.02 ± 0.00 d | 0.089 ± 0.004 c | 0.02 ± 0.00 d |
| 34 | Quercetin-3-rhamnoside | nd | nd | nd | nd | 2.26 ± 0.38 b | 4.31 ± 0.51 a | nd |
| 35 | Quercetin-3-xyloside | 0.01 ± 0.00 e | 0.02 ± 0.00 de | 0.06 ± 0.00 c | 0.56 ± 0.04 a | 0.09 ± 0.01 b | 0.05 ± 0.01 cd | 0.02 ± 0.00 de |
| 36 | Quercetin-arabinofuranoside | 0.13 ± 0.04 c | 0.15 ± 0.02 c | 0.25 ± 0.03 b | 0.17 ± 0.00 c | 0.03 ± 0.01 d | 0.31 ± 0.04 a | 0.004 ± 0.000 d |
| 37 | Quercetin-arabinopyranoside | 0.01 ± 0.00 c | nd | nd | nd | 2.07 ± 0.20 a | 1.61 ± 0.05 b | nd |
| 38 | Quercetin-acetyl hexoside 1 | nd | 4.00 ± 0.40 a | 1.53 ± 0.07 b | 0.22 ± 0.00 c | nd | nd | nd |
| 39 | Quercetin-acetyl hexoside 2 | nd | 0.14 ± 0.02 b | 0.08 ± 0.00 c | 0.35 ± 0.02 a | nd | nd | nd |
| 40 | Kaempferol trihexoside | nd | 1.27 ± 0.11 a | 0.81 ± 0.08 b | nd | nd | nd | nd |
| 41 | Kaempferol-3-rutinoside | 0.92 ± 0.02 a | 0.06 ± 0.00 f | 0.05 ± 0.01 f | 0.54 ± 0.01 b | 0.37 ± 0.08 c | 0.24 ± 0.03 d | 0.15 ± 0.01 e |
| 42 | Kaempferol acetyl hexoside 1 | nd | 1.50 ± 0.15 a | 0.32 ± 0.01 b | 0.09 ± 0.01 a | nd | nd | nd |
| 43 | Kaempferol acetyl hexoside 2 | nd | 0.07 ± 0.01 a | nd | 0.33 ± 0.04 a | nd | nd | nd |
| 44 | Kaempferol dihexoside | nd | 0.12 ± 0.01 b | 0.44 ± 0.03 a | nd | nd | nd | nd |
| 45 | Kaempferol pentoside 1 | nd | nd | nd | 1.34 ± 0.09 a | nd | nd | nd |
| 46 | Kaempferol pentoside 2 | nd | nd | nd | 0.12 ± 0.01 a | nd | nd | nd |
| 47 | Kaempferol rhamnoside | 0.06 ± 0.01 b | nd | nd | nd | 0.32 ± 0.08 b | 4.43 ± 0.63 a | nd |
| 48 | Kaempferol hexoside 1 | 0.01 ± 0.00 e | 0.20 ± 0.02 b | 0.08 ± 0.00 d | 0.27 ± 0.02 a | 0.03 ± 0.01 e | 0.02 ± 0.00 e | 0.14 ± 0.01 c |
| 49 | Kaempferol hexoside 2 | 0.69 ± 0.23 a | nd | nd | 0.38 ± 0.01 b | 0.01 ± 0.00 c | 0.34 ± 0.04 b | nd |
| 50 | Kaempferol rhamnosyl hexoside | nd | nd | nd | nd | 0.02 ± 0.01 b | 0.22 ± 0.03 a | nd |
| 51 | Isorhamnetin hexoside | nd | 0.01 ± 0.00 c | 0.04 ± 0.00 c | 0.04 ± 0.00 c | 0.28 ± 0.04 b | 3.12 ± 0.21 a | 0.22 ± 0.00 b |
| 52 | Isorhamnetin dihexoside | 0.31 ± 0.02 a | nd | nd | nd | nd | nd | nd |
| 53 | Isorhamnetin acetyl hexoside 1 | nd | nd | nd | nd | nd | nd | 0.98 ± 0.12 a |
| 54 | Isorhamnetin acetyl hexoside 2 | nd | nd | nd | nd | nd | nd | 0.04 ± 0.00 a |
| 55 | Isorhamnetin-3-rutinoside | 0.02 ± 0.00 a | 0.01 ± 0.00 c | 0.02 ± 0.00 b | 0.02 ± 0.00 b | nd | nd | nd |
| 56 | Myricetin rutinoside | nd | nd | nd | nd | nd | nd | 0.004 ± 0.000 a |
| 57 | Laricitrin glucuronide | 0.07 ± 0.01 a | nd | nd | nd | nd | nd | nd |
| 58 | Syringetin hexoside 1 | 0.02 ± 0.00 b | nd | nd | nd | 0.30 ± 0.03 b | 5.18 ± 0.62 a | nd |
| 59 | Syringetin hexoside 2 | 0.29 ± 0.10 a | nd | nd | nd | nd | nd | nd |
| 60 | Syringetin acetyl hexoside 1 | nd | nd | nd | nd | 1.61 ± 0.41 a | 0.78 ± 0.14 b | 0.30 ± 0.02 b |
| 61 | Syringetin acetyl hexoside 2 | nd | nd | nd | nd | nd | nd | 0.07 ± 0.00 a |
| Total identified flavonols | 5.93 ± 0.05 b | 8.71 ± 0.06 b | 9.42 ± 0.03 b | 8.37 ± 0.02 b | 9.29 ± 0.11 b | 21.60 ± 0.16 a | 2.84 ± 0.02 c | |
| 62 | Apigenin hexoside | nd | 0.02 ± 0.00 b | 0.01 ± 0.00 b | 0.04 ± 0.00 a | nd | nd | nd |
| Total identified flavones | nd | 0.02 ± 0.00 b | 0.01 ± 0.00 b | 0.04 ± 0.00 a | nd | nd | nd | |
| 63 | Phloretin xylosylglucoside | nd | nd | nd | nd | 0.18 ± 0.03 a | 0.09 ± 0.02 b | nd |
| 64 | Phloridzin | nd | nd | nd | nd | 5.14 ± 0.74 a | 5.23 ± 0.52 a | nd |
| 65 | Trilobatin | nd | nd | nd | nd | 0.30 ± 0.08 b | 1.47 ± 0.20 a | nd |
| Total identified chalcones | nd | nd | nd | nd | 5.61 ± 0.28 a | 6.80 ± 0.24 a | nd | |
| 66 | Procyanidin dimer 1 | nd | 0.33 ± 0.03 c | 0.52 ± 0.03 c | 1.24 ± 0.04 c | 1.45 ± 0.42 bc | 2.52 ± 0.36 b | 7.90 ± 1.48 a |
| 67 | Procyanidin dimer 2 | nd | nd | nd | 5.07 ± 0.19 a | 0.75 ± 0.31 c | 2.93 ± 0.47 b | 5.30 ± 0.27 a |
| 68 | Procyanidin dimer 3 | nd | nd | nd | nd | 0.02 ± 0.01 b | 3.43 ± 0.40 a | 0.36 ± 0.02 b |
| 69 | Procyanidin dimer 4 | nd | nd | nd | nd | 0.03 ± 0.01 c | 3.43 ± 0.39 b | 6.31 ± 0.25 a |
| 70 | Procyanidin dimer 5 | nd | nd | nd | nd | nd | nd | 1.21 ± 0.08 a |
| 71 | Procyanidin dimer 6 | nd | nd | nd | nd | nd | nd | 0.50 ± 0.23 a |
| 72 | Procyanidin trimer 1 | nd | 1.18 ± 0.09 d | 0.86 ± 0.05 d | 11.05 ± 0.30 a | 2.15 ± 0.31 c | 4.76 ± 0.39 b | 0.02 ± 0.00 e |
| 73 | Procyanidin trimer 2 | nd | 2.63 ± 0.18 c | 2.68 ± 0.16 c | 5.97 ± 0.55 a | 0.12 ± 0.05 d | 0.07 ± 0.01 d | 4.52 ± 0.16 b |
| 74 | Procyanidin trimer 3 | nd | nd | nd | nd | 0.42 ± 0.04 b | 8.66 ± 1.08 a | 0.02 ± 0.00 b |
| 75 | Procyanidin trimer 4 | nd | nd | nd | nd | 2.33 ± 0.58 a | 1.72 ± 0.28 a | nd |
| 76 | Procyanidin trimer 5 | nd | nd | nd | nd | nd | 4.10 ± 1.60 a | nd |
| 77 | Procyanidin tetramer | nd | nd | nd | nd | nd | 4.11 ± 1.60 a | 2.89 ± 0.18 a |
| Total identified condensed tannins | nd | 4.14 ± 0.10 c | 4.06 ± 0.08 c | 23.33 ± 0.27 b | 7.27 ± 0.22 c | 35.73 ± 0.66 a | 29.03 ± 0.27 b | |
| Total identified compounds | 16.44 ± 0.06 d | 22.05 ± 0.07 d | 41.54 ± 0.06 c | 65.10 ± 0.09 b | 27.62 ± 0.14 d | 82.77 ± 0.26 a | 46.49 ± 0.09 c |
3. LC-DAD-MS Analysis of the Individual Phenolics in the Inflorescences
4. Antidiabetic Activity of the Inflorescences
| TP | TF | TNF | TT | CT | SS | ABTS | FRAP | DPPH | HepG2 | HCT116 | HaCaT | α-Amyl | α-Glucos | 5-Lipoxy | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TP | 1.000 | ||||||||||||||
| TF | 0.761 | 1.000 | |||||||||||||
| TNF | 0.938 | 0.629 | 1.000 | ||||||||||||
| TT | 0.471 | 0.014 | 0.333 | 1.000 | |||||||||||
| CT | −0.065 | 0.178 | −0.367 | 0.314 | 1.000 | ||||||||||
| SS | 0.194 | −0.210 | 0.251 | 0.262 | 0.045 | 1.000 | |||||||||
| ABTS | 0.937 | 0.875 | 0.908 | 0.160 | −0.140 | 0.115 | 1.000 | ||||||||
| FRAP | 0.873 | 0.913 | 0.862 | 0.016 | −0.176 | −0.022 | 0.979 | 1.000 | |||||||
| DPPH | 0.830 | 0.952 | 0.762 | −0.015 | −0.094 | −0.170 | 0.945 | 0.969 | 1.000 | ||||||
| HepG2 | −0.459 | −0.567 | −0.396 | −0.336 | −0.353 | 0.059 | −0.451 | −0.489 | −0.393 | 1.000 | |||||
| HCT116 | −0.416 | −0.353 | −0.327 | −0.602 | −0.171 | 0.454 | −0.250 | −0.274 | −0.265 | 0.689 | 1.000 | ||||
| HaCaT | −0.148 | −0.067 | −0.029 | −0.670 | −0.512 | −0.059 | −0.004 | 0.050 | 0.104 | 0.729 | 0.623 | 1.000 | |||
| α-amyl | 0.169 | 0.458 | −0.116 | −0.038 | 0.585 | −0.064 | 0.237 | 0.191 | 0.377 | 0.146 | 0.234 | 0.117 | 1.000 | ||
| α-glucos | 0.077 | −0.083 | −0.101 | 0.645 | 0.627 | 0.429 | −0.065 | −0.205 | −0.213 | −0.312 | −0.121 | −0.786 | 0.263 | 1.000 | |
| 5-lipoxy | −0.345 | 0.020 | −0.371 | −0.530 | 0.431 | 0.318 | −0.172 | −0.114 | −0.152 | −0.077 | 0.534 | 0.154 | 0.232 | 0.011 | 1.000 |
5. Anti-Inflammatory Activity of Inflorescences
6. Cytotoxic Activity of the Inflorescences
| Cell Type (IC50 μg/mL) | |||
|---|---|---|---|
| HepG2 | HCT 116 | HaCaT | |
| Prunus avium | 300.89 ± 0.21 c A | 261.97 ± 13.12 c A | 323.84 ± 46.61 c A |
| Prunus serrulata | 473.59 ± 35.69 ab A | 517.42 ± 37.10 a A | 377.66 ± 34.85 bc B |
| Prunus serrulata 2018Kiku Shidare Zakura’ | 409.71 ± 103.52 b A | 464.01 ± 57.31 a A | 385.20 ± 7.27 bc A |
| Prunus yedoensis | 508.09 ± 26.28 a A | 537.92 ± 43.0 a A | 521.64 ± 67.29 a A |
| Malus purpurea | 386.2 ± 19.92 b B | 539.66 ± 45.19 a A | 461.39 ± 71.56 ab AB |
| Malus floribunda | 445.78 ± 27.42 ab A | 361.83 ± 31.19 b B | 459.28 ± 43.69 ab A |
| Chaenomeles japonica | 452.48 ± 15.18 ab A | 470.66 ± 48.16 a A | 473.27 ± 92.54 ab A |
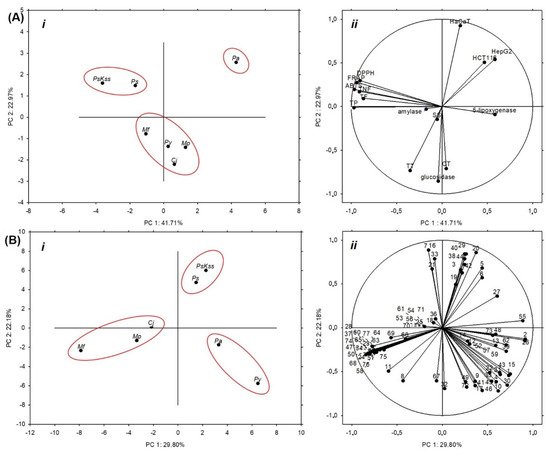
7. Statistical Analysis
This entry is adapted from the peer-reviewed paper 10.3390/plants11030271
References
- Orlando, G.; Recinella, L.; Chiavaroli, A.; Brunetti, L.; Leone, S.; Carradori, S.; Di Simone, S.; Ciferri, M.C.; Zengin, G.; Ak, G.; et al. Water extract from inflorescences of industrial hemp futura 75 variety as a source of anti-inflammatory, anti-proliferative and antimycotic agents: Results from in silico, in vitro and ex vivo studies. Antioxidants 2020, 9, 437.
- Lau, B.F.; Kong, K.W.; Leong, K.H.; Sun, J.; He, X.; Wang, Z.; Mustafa, M.R.; Ling, T.C.; Ismail, A. Banana inflorescence: Its bio-prospects as an ingredient for functional foods. Trends Food Sci. Technol. 2020, 97, 14–28.
- Sarv, V.; Venskutonis, P.R.; Bhat, R. The Sorbus spp.—Underutilised plants for foods and nutraceuticals: Review on polyphenolic phytochemicals and antioxidant potential. Antioxidants 2020, 9, 813.
- El Euch, S.K.; Bouajila, J.; Bouzouita, N. Chemical composition, biological and cytotoxic activities of Cistus salviifolius flower buds and leaves extracts. Ind. Crops Prod. 2015, 76, 1100–1105.
- Lee, E.J.; Kim, J.S.; Kim, H.P.; Lee, L.-H.; Kang, S.S. Phenolic constituents from the flower buds of Lonicera japonica and their 5-lipoxygenase inhibitory activities. Food Chem. 2010, 120, 134–139.
- Ferreres, F.; Andrade, C.; Gomes, N.G.; Andrade, P.B.; Gil-Izquierdo, A.; Pereira, D.M.; Suksungworn, R.; Duangsrisai, S.; Videira, R.A.; Valentão, P. Valorisation of kitul, an overlooked food plant: Phenolic profiling of fruits and inflorescences and assessment of their effects on diabetes-related targets. Food Chem. 2021, 342, 128323.
- Olszewska, M.A.; Michel, P. Antioxidant activity of inflorescences, leaves and fruits of three Sorbus species in relation to their polyphenolic composition. Nat. Prod. Res. 2009, 23, 1507–1521.
- Jesus, F.; Gonçalves, A.C.; Alves, G.; Silva, L.R. Health benefits of Prunus avium plant parts: An unexplored source rich in phenolic compounds. Food Rev. Int. 2020.
- Acero, N.; Gradillas, A.; Beltran, M.; García, A.; Muñoz Mingarro, D. Comparison of phenolic compounds profile and antioxidant properties of different sweet cherry (Prunus avium L.) varieties. Food Chem. 2019, 279, 260–271.
- Usenik, V.; Fabčič, J.; Štampar, F. Sugars, organic acids, phenolic composition and antioxidant activity of sweet cherry (Prunus avium L.). Food Chem. 2008, 107, 185–192.
- Kang, G.-J.; Lee, H.-J.; Yoon, W.-J.; Yang, E.-J.; Park, S.-S.; Kang, H.-K.; Park, M.-H.; Yoo, E.-S. Prunus yedoensis inhibits the inflammatory chemokines, MDC and TARC, by regulating the STAT1-signaling pathway in IFN-γ-stimulated HaCaT human keratinocytes. Biomol. Ther. 2008, 16, 394–402.
- Teleszko, M.; Wojdyło, A. Comparison of phenolic compounds and antioxidant potential between selected edible fruits and their leaves. J. Funct. Foods 2015, 14, 736–746.
- Zhang, X.; Su, M.; Du, J.; Zhou, H.; Li, X.; Li, X.; Ye, Z. Comparison of phytochemical differences of the pulp of different peach (Prunus persica (L.) Batsch) cultivars with alpha-glucosidase inhibitory activity variations in China using UPLC-Q-TOF/MS. Molecules 2019, 24, 1968.
- Wojdyło, A.; Nowicka, P.; Grimalt, M.; Legua, P.; Almansa, M.S.; Amorós, A.; Carbonell-Barrachina, A.; Hernández, F. Polyphenol compounds and biological activity of caper (Capparis spinosa L.) flowers buds. Plants 2019, 8, 539.
- Borneo, R.; León, A.E.; Aguirre, A.; Ribotta, P.; Cantero, J.J. Antioxidant capacity of medicinal plants from the Province of Córdoba (Argentina) and their in vitro testing in a model food system. Food Chem. 2009, 112, 664–670.
- Afonso, S.; Oliveira, I.V.; Meyer, A.S.; Aires, A.; Saavedra, M.J.; Gonçalves, B. Phenolic profile and bioactive potential of stems and seed kernels of sweet cherry fruit. Antioxidants 2020, 9, 1295.
- Yang, S.; Meng, Z.; Li, Y.; Chen, R.; Yang, Y.; Zhao, Z. Evaluation of physiological characteristics, soluble sugars, organic acids and volatile compounds in ‘Orin’ apples (Malus domestica) at different ripening stages. Molecules 2021, 26, 807.
- Walker, R.P.; Battistelli, A.; Bonghi, C.; Drincovich, M.F.; Falchi, R.; Lara, M.V.; Moscatello, S.; Vizzotto, G.; Famiani, F. Non-structural carbohydrate metabolism in the flesh of stone fruits of the genus Prunus (Rosaceae)—A review. Front. Plant Sci. 2020, 11, 549921.
- Bastos, C.; Barros, L.; Dueñas, M.; Calhelha, R.C.; Queiroz, M.J.R.; Santos-Buelga, C.; Ferreira, I.C. Chemical characterisation and bioactive properties of Prunus avium L.: The widely studied fruits and the unexplored stems. Food Chem. 2015, 173, 1045–1053.
- Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P.; Golis, T.; Bąbelewski, P. ABTS On-line antioxidant, α-amylase, α-glucosidase, pancreatic lipase, acetyl-and butyrylcholinesterase inhibition activity of Chaenomeles fruits determined by polyphenols and other chemical compounds. Antioxidants 2020, 9, 60.
- Fang, L.; Meng, W.; Min, W. Phenolic compounds and antioxidant activities of flowers, leaves and fruits of five crabapple cultivars (Malus Mill. species). Sci. Hortic. 2018, 235, 460–467.
- Gonçalves, A.C.; Bento, C.; Silva, B.M.; Silva, L.R. Sweet cherries from Fundão possess antidiabetic potential and protect human erythrocytes against oxidative damage. Food Res. Int. 2017, 95, 91–100.
- Gonçalves, B.; Landbo, A.K.; Knudsen, D.; Silva, A.P.; Moutinho-Pereira, J.; Rosa, E.; Meyer, A.S. Effect of ripeness and postharvest storage on the phenolic profiles of cherries (Prunus avium L.). J. Agric. Food Chem. 2004, 52, 523–530.
- Olszewska, M. Quantitative HPLC analysis of flavonoids and chlorogenic acid in the leaves and inflorescences of Prunus serotina Ehrh. Acta Chromatogr. 2007, 19, 253–269.
- Olszewska, M.A.; Kwapisz, A. Metabolite profiling and antioxidant activity of Prunus padus L. flowers and leaves. Nat. Prod. Res. 2011, 25, 1115–1131.
- Coklar, H.; Akbulut, M.; Alhassan, I.; Kirpitci, Ş.; Korkmaz, E. Organic acids, sugars, phenolic compounds and antioxidant activity of Malus floribunda coccinella fruit, peel and flesh. Acta Sci. Pol. Hortorum Cultus 2018, 17, 47–59.
- Du, H.; Wu, J.; Li, H.; Zhong, P.-X.; Xu, Y.-J.; Li, C.-H.; Ji, K.-X.; Wang, L.-S. Polyphenols and triterpenes from Chaenomeles fruits: Chemical analysis and antioxidant activities assessment. Food Chem. 2013, 141, 4260–4268.
- Valcheva-Kuzmanova, S.V.; Denev, P.N.; Ognyanov, M.H. Chemical composition and antioxidant activity of Chaenomeles maulei fruit juice. J. Biomed. Clin. Res. 2018, 11, 41–48.
- Da Silva, S.M.; Koehnlein, E.A.; Bracht, A.; Castoldi, R.; de Morais, G.R.; Baesso, M.L.; Peralta, R.; de Souza, C.G.M.; de Sá-Nakanishi, A.B.; Peralta, R.M. Inhibition of salivary and pancreatic α-amylases by a pinhão coat (Araucaria angustifolia) extract rich in condensed tannin. Food Res. Int. 2014, 56, 1–8.
- Mkandawire, N.L.; Kaufman, R.C.; Bean, S.R.; Weller, C.L.; Jackson, D.S.; Rose, D.J. Effects of sorghum (Sorghum bicolor (L.) Moench) tannins on alpha-amylase activity and in vitro digestibility of starch in raw and processed flours. J. Agric. Food Chem. 2013, 61, 4448–4454.
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2021, 338, 128119.
- Assefa, S.T.; Yang, E.Y.; Chae, S.Y.; Song, M.; Lee, J.; Cho, M.C.; Jang, S. Alpha glucosidase inhibitory activities of plants with focus on common vegetables. Plants 2020, 9, 2.
- Kumar, V.; Sachan, R.; Rahman, M.; Sharma, K.; Al-Abassi, F.A.; Anwar, F. Prunus amygdalus extract exert antidiabetic effect via inhibition of DPP-IV: In-silico and in-vivo approaches. J. Biomol. Struct. Dyn. 2021, 39, 4160–4174.
- Meier, J.J. GLP-1 receptor agonists for individualized treatment of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2012, 8, 728–742.
- Seino, Y.; Fukushima, M.; Yabe, D. GIP and GLP-1, the two incretin hormones: Similarities and differences. J. Diabetes Investig. 2010, 1, 8–23.
- Wang, T.K.; Xu, S.; Li, S.; Zhang, Y. Proanthocyanidins should be a candidate in the treatment of cancer, cardiovascular diseases and lipid metabolic disorder. Molecules 2020, 25, 5971.
- Xu, Y.Q.; Gao, Y.; Granato, D. Effects of epigallocatechin gallate, epigallocatechin and epicatechin gallate on the chemical and cell-based antioxidant activity, sensory properties, and cytotoxicity of a catechin-free model beverage. Food Chem. 2021, 339, 128060.
- Muccilli, V.; Cardullo, N.; Spatafora, C.; Cunsolo, V.; Tringali, C. α-Glucosidase inhibition and antioxidant activity of an oenological commercial tannin. Extraction, fractionation and analysis by HPLC/ESI-MS/MS and 1H NMR. Food Chem. 2017, 215, 50–60.