Expression and functions of microRNAs (miRNAs) have been widely investigated in cancer treatment-induced complications and as a response to physical activity, respectively, but few studies focus on the application of miRNAs as biomarkers in exercise-based cancer rehabilitation. Research has shown that certain miRNA expression is altered substantially due to tissue damage caused by cancer treatment and chronic inflammation. MiRNAs are released from the damaged tissue and can be easily detected in blood plasma. Levels of the miRNA present in peripheral circulation can therefore be used to measure the extent of tissue damage. Moreover, damage to tissues such as cardiac and skeletal muscle significantly affects the individual’s health-related fitness, which can be determined using physiologic functional assessments. These physiologic parameters are a measure of tissue health and function and can therefore be correlated with the levels of circulating miRNAs.
1. Introduction to miRNAs
miRNAs are short, 20-22-nucleotide, noncoding RNAs that inhibit the expression of their cognate target genes by directly binding to their mRNAs, thus blocking their translation and targeting them for degradation. In this way, miRNAs play an essential part in regulating gene expression in cells. Various studies have elucidated that the aberrant expression of miRNAs is associated with the onset and progress of multiple diseases, including cancer, diabetes, inflammation, cardiovascular disease, neurological disorders, etc. Evidence has also shown that miRNAs can act as intracellular modulators of responses to cancer treatment and treatment-induced complications or toxicities
[1][2]. MiRNAs have been attributed with several characteristics such as (1) presence in multiple body fluids, including blood; (2) high stability; (3) resistance to RNA enzyme, thus resulting in long half-life; (4) tissue-specific function; and (5) easy detection and quantitation by different methods. These properties make miRNAs amenable to a wide range of applications in diagnosis, prognosis, and treatment in multiple disease settings.
Cells constantly secrete small vesicles (30–150 nm) called exosomes, which are found in abundance in all body fluids including blood. These vesicles contain RNA and protein secreted from the cells from which the exosome originates and are involved in many biological functions, including cell-to-cell communication and signalling
[3][4]. Since exosomes represent a snapshot of the cellular contents, any condition that affects the normal functions of the cell is likely to alter the composition of exosomes. As a result, RNA and protein present in the exosomes can be used as biomarkers that can have both prognostic and diagnostic applications
[3][4]. Consequently, profiling of circulating RNAs, in particular miRNAs, have been used in a number of studies to identify novel and highly promising biomarkers for many pathologies including neurodegenerative diseases such as Alzheimer’s and Parkinson’s, brain injury, presence of brain tumours, and in cancer (breast and pancreatic cancers)
[3][4]. Many of these molecules were determined to be associated with exosomes. Exosomes can be easily purified and analysed and therefore have gained importance in research as a source for identification of novel biomarkers.
Although miRNAs have been widely studied in diseases and in response to physical activity (as shown in Figure 1), few studies have investigated the application of miRNAs as a dynamic monitoring tool during exercise-based cancer rehabilitation.
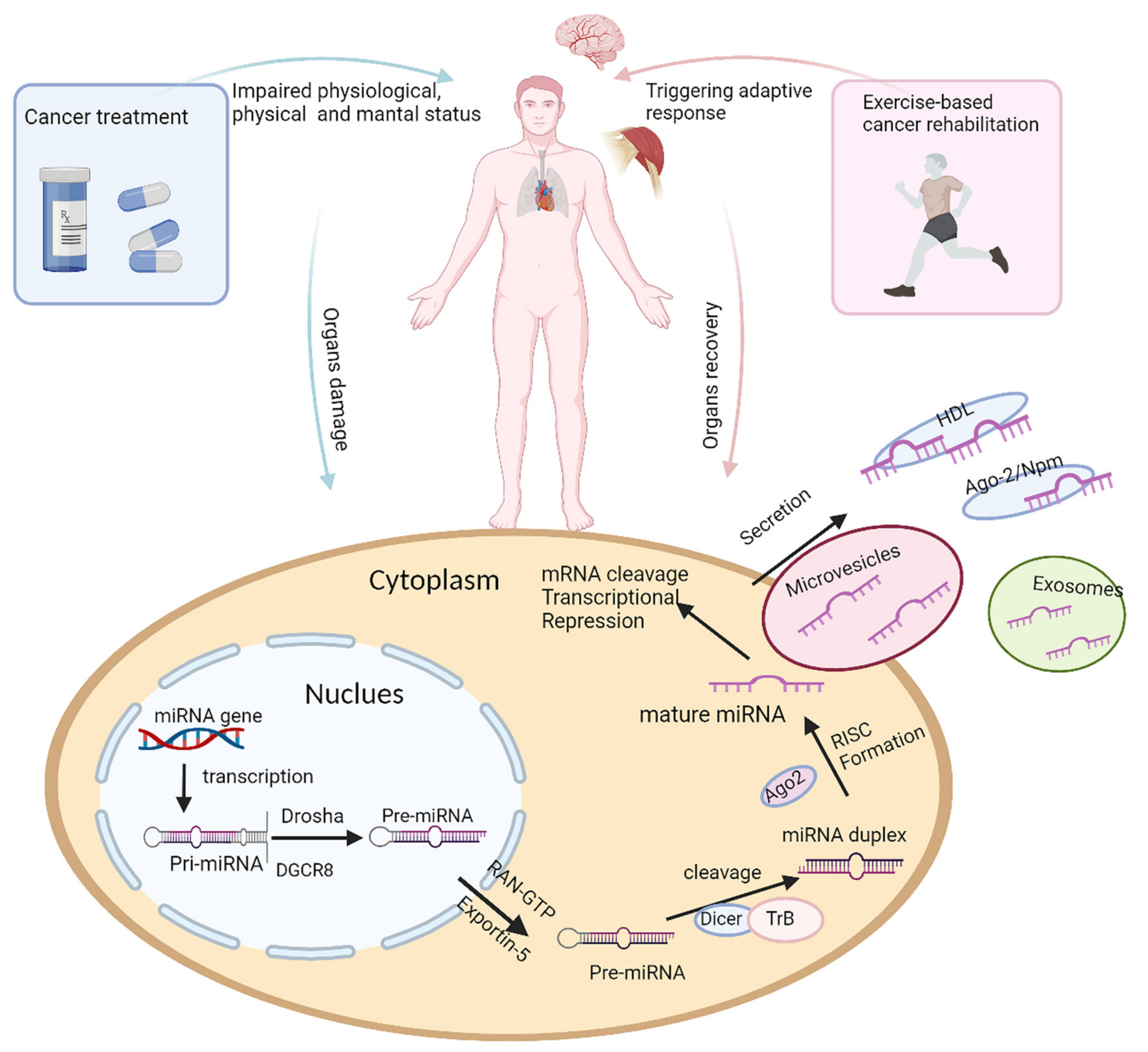
Figure 1. The impact of cancer treatment and physical activity on miRNA biogenesis. Organ damage can be caused as a result of cancer or as an adverse effect of cancer treatment, leading to decreased physical and mental ability and poor quality of life for cancer survivors, while exercise-based cancer rehabilitation triggers adaptive responses, resulting in altered miRNA expression, in damaged organs, thus promoting in organ recovery and improved quality of life. The diagram illustrates the changes in miRNA expression that can occur during tissue damage, toxicity, treatment side effects, and during physical activity, and this is part of the adaptive process or physiologic responses to exercise. During tissue damage, miRNAs may be released into circulation. miRNA biogenesis and molecular mechanisms of miRNA production can also be altered by disease or as a response to physical activity. miRNA biogenesis begins in the nucleus, where primary miRNA (pri-miRNA), transcribed by miRNA gene, is processed by Drosha and Dgcr8 into stem-looped structures, precursor miRNA (pre-miRNA). Pre-miRNA is exported by Exportin 5 into cytoplasm, where mature miRNA is produced after further processing by Dicer. Mature miRNA is transported out of cell by either exocytosis or combining protein complex, regulating gene expression in cancer treatment-induced complications or adaptive changes during exercise-based cancer rehabilitation.
2. Potential miRNAs as Biomarkers in Exercise-Based Cancer Rehabilitation
2.1. Metabolic Syndrome
2.1.1. miR-126 and miR-146a
Gomes et al.
[5] showed that obesity could lead to decreased expression of miR-126 and an increased
PI3KR2, which inhibits a key regulator of the
VEGF signalling pathway
PI3K, thus causing capillary rarefaction in skeletal muscle in the obese rat model, whereas aerobic exercise training, such as swimming, can reverse this negative impact by normalizing miR-126 level and restoring
VEGF signalling. Of note, other studies showed miR-126 increased in obese patients, but after physical activity, there was a slight decrease of miR-126, without statistical significance
[6].
miRNA was associated with the metabolic disorder by regulating the inflammatory response in white adipose tissue, leading to insulin resistance. miR-146a can suppress the macrophage-conditioned medium-induced inflammatory response in adipocytes, which was reflected by a decrease of
IL-8, MCP-1 mRNA and protein. The mechanism of this process involves a reduced inflammation-induced activation of
JNK and
p38 via targeting
IRAK1 and
TRAF6 in human adipocytes
[7]. Studies
[6] showed that miR-146a-5p significantly increased in obese patients. The increase of miR-146a-5p was also accompanied by an increase of inflammatory genes, such as
TRL4, NF-kB, IL-6, and
TNF-α in human mononuclear leukocytes, while, after exposure to physical activity, four-fifths of responders in these obese patients had a remarkable decrease of miR-146a-5p. In addition, miR-146a-5p was significantly correlated to lipid parameters, namely total cholesterol and inflammatory cytokine
IL-8. These results indicated that miR-146a-5p might play a critical role in obesity by activating the inflammatory response and serve as a promising biomarker for obesity and its metabolic complications during exercise.
2.1.2. miR-9 and miR-375
Studies showed miR-375 and miR-9 presented a positive correlation to glycaemic status. Moreover, miR-375 showed good diagnostic abilities to distinguish prediabetes and T2DM patients from healthy controls, with area under curve (AUC) of 0.76 (95% confidence interval (CI): 0.630–0.884,
p = 0.001) and 0.77 (95% CI: 0.65–0.89,
p < 0.001) compared to miR-9 (prediabetes: AUC = 0.63, 95% CI: 0.485–0.777,
p = 0.08; T2DM: AUC = 0.50, 95% CI: 0.301–0.604,
p = 0.053). Results also showed that the combination of miR-375 and miR-9 significantly enhanced the predictability to distinguish patients from controls. Taken together, miR-375 alone or combined with miR-9 could be used as biomarkers for prediabetes and T2DM
[8]. Inhibiting insulin exocytosis by miR-9 and reducing the number and viability of pancreatic β-cells by overexpression of miR-375 may provide a theoretical basis for their application in T2DM diagnosis. Despite the diagnostic value or valid theoretical basis, few studies investigate the change of these two miRNAs in metabolic syndrome during exercise in cancer survivors.
2.1.3. Others
Other miRNAs, such as miR-92a, miR-130a, miR-222, and miR-370, were remarkably decreased in pre-atherosclerotic patients (patients without atherosclerosis but having hypertension, hyperlipidaemia, and/or diabetes), with miR-126 and miR-130a decreased up to 50%
[9]. Like miR-375 and miR-9, these miRNAs had been deeply investigated for the mechanism of the initiation of metabolic disease. Still, these studies did not establish a clinical correlation between metabolic syndrome and the miRNAs, and there was no overlap identified with physical activity or exercise training. Therefore, whether these miRNAs can be used for potential biomarkers of exercise-based cancer rehabilitation remains to be determined.
2.2. Cardiorespiratory Capacity
2.2.1. myomiRs
The elevation of circulating miR-1 was significantly associated with LVEF reduction, and the receiver operating characteristic curve (ROC) analysis showed that miR-1 exceeded cardiac troponin I (cTnI) in distinguishing potential victims to cardiotoxicity
[10]. In rat models, 14 weeks of endurance training significantly increased the expression of miR-1 and miR-133, which can result in physiological hypertrophy by targeting downstream
Srf, Hdac4, and
Hand2 gene
[11]. Moreover, circulating miR-1, miR-133a, and miR-206 were correlated to aerobic performance parameters such as VO
2max and running speed at individual anaerobic lactate thresholds in marathon runners
[12]. Nielsen et al.
[13] demonstrated that the expression of myomiRs in muscle biopsies, such as miR-1, miR-133a, miR-133b, and miR-206, were related to increased VO
2max and improved insulin sensitivity after endurance training. These results suggested that these myomiRs play a pivotal role in physical adaption during exercise. Nevertheless, Wardle et al.
[14] found several myomiRs (miR-1, miR-133a, miR-206, miR-499) expressed at very low levels in the plasma. Although miRNAs at a low level are sufficient to cause physiological changes, whether or not the low level of miRNAs impacts, the correlation analysis remains unknown.
miR-208a, miR-208b, and miR-499 are all encoded by the myosin gene in muscle. Although miR-208a is cardiac-specific and its role in doxorubicin-induced cardiotoxicity has been validated in the mice model, its role in cancer patients seemed disappointing
[10][15]. miR-208b and miR-499 were reported to rebound back to pre-exercise levels after 24h of completion of the exercise in marathon runners. They were not correlated with cardiac injury markers, such as cTnI and B-type natriuretic peptide (NT-pro-BNP)
[12]. These results suggested the limited role of these miRNAs to evaluate or monitor cardiovascular capacity in physical activity.
2.2.2. miR-126
In rat models, miR-126, as a pre-angiogenic miRNA, played a critical role in having a cardioprotective effect after exposure to crocin and voluntary exercise, the mechanism of which involved facilitated cardiac angiogenesis through
Akt and
ERK1/2 pathways
[6]. Furthermore, miR-126, which HIF-1α induced during exercise training, also contributed to myocardial angiogenesis via the
PI3K/AKT/eNOS and
MAPK signalling pathway and subsequently improved heart function in myocardial infarction rats
[16]. Exosomes from miR-126 overexpressing mesenchymal stem cells were shown to induce angiogenesis via the
PIK3R2-mediated
PI3K/Akt signalling pathway
[7]. Moreover, miR-126 was positively correlated to NT-pro-BNP and cTN-I in atherosclerotic patients
[9]. Exercise training reduced the high-density lipoprotein (HDL)-induced miR-126 in patients with chronic heart failure and eventually caused atherogenesis and endothelial dysfunction
[17]. These results suggested miR-126 was associated with common indicators of cardiac damage and may serve as a potential biomarker for treatment-induced cardiotoxicity as well we exercise-based cardiac and cancer rehabilitation.
2.2.3. miR-21 and miR-146a
miR-21 and miR-146a are highly responsive to both cardiac damage and physical activity. miR-21 can attenuate diastolic dysfunction of diabetic cardiomyopathy via targeting gelsolin
[18]. Like miR-126, miR-21, as a mediator, can promote hypoxia-induced angiogenesis in the cardiac microvascular endothelial cells
[19]. Other studies have shown the relationship between miRNAs and common indicators of cardiac damage. miR-21 was negatively correlated to NT-pro-BNP and cTN-I among atherosclerotic patients
[9]. In myocardial infarction in rat models, the injection of miR-146 antagomiR improved the reduction of left ventricular ejection fraction LVEF and fractional shortening. miR-146 antagomiR also reduced the levels of atrial natriuretic peptide (ANP) and BNP mRNA. In addition, miR-146 myomiRs attenuated cardiac fibrosis by reducing the increase of collagen I and collagen III mRNA. These results demonstrated that inhibition of miR-146 ameliorated cardiac dysfunction and remodelling
[20]. A linear correlation between circulating miR-146a and cardiac marker creatine kinase-MB (CKMB) isoenzyme as well as high sensitivity C-reactive protein (hs-CRP) was observed in basketball players
[21]. Studies showed that miR-21 and miR-146a had an opposite expression pattern and function during exercise: miR-21 had proinflammatory effects and was upregulated after acute exercise but downregulated after endurance training. By contrast, miR-146a had a proinflammatory effect and was upregulated by acute cycling exercise before and after 90 days of sustained rowing training. Additionally, the expression of miR-21 was negatively correlated with VO
2max, while miR-146a was positively correlated with VO
2max
[22][23], suggesting promising prospects in an exercise-based rehabilitation program.
2.2.4. miR-222
In a study with
HER-2 positive breast cancer patients undergoing neoadjuvant target therapy Trastuzumab, serum miR-222-3p protected against the relative drop of LVEF (OR = 0.410, 95% CI: 0.175–0.962,
p = 0.040) and absolute drop of LVEF (OR = 0.394, 95% CI: 0.166–0.937,
p = 0.035), demonstrating that miR-222-3p was an independent protective factor for trastuzumab-induced cardiotoxicity
[24]. Recent work suggested that high-intensity interval training increased miR-222 expression after an acute bout of exercise and following a sustained period of exercise
[25]. Liu et al.
[26] suggested that miR-222 is necessary for exercise-induced cardiac growth and to counteract pathological cardiac remodelling via targeting
P27, HIPK1, and
HMBOX1. This may explain how exercise benefits cardiovascular fitness. Despite the positive effects of miR-222 on cardiac physiology, miR-222 can inhibit
p27Kip1, a cyclin-dependent kinase inhibitor, thus promoting cancer growth
[27]. Moreover, a low level of serum miR-222 was associated with superior pathological complete response, disease-free survival, and overall survival in breast cancer patients who received Trastuzumab
[24]. These results suggested that miR-222 was a promising biomarker for cardiac fitness but a double-edged sword in the QoL of cancer survivors.
2.2.5. miR-155
In fibrotic illnesses, miR-155 is persistently elevated, and knocking it out reduces collagen synthesis
[28]. In vivo studies showed that miR-155-5p was significantly downregulated in radiation-induced EMT cell model, while in vitro studies showed that ectopic overexpression of miRNA-155-5p inhibited EMT through the NF-KB pathway by targeting
GSK-3β. This alleviated radiation-induced pulmonary fibrosis
[29]. In the mouse model of bleomycin-induced pulmonary fibrosis, the
miRNA–155/LXR pathway was identified to play an important role in experimental and idiopathic pulmonary fibrosis
[30]. In another study, untrained rats showed enhanced acetylcholine (Ach)-induced relaxation and nitric oxide (NO) production compared to trained rats. Results showed a higher expression of endothelial NO synthase (eNOS), the posttranslational modification of which by the
PI3-kinase/Akt1/2/3 pathway, contributed to higher production of exercise-induced NO expression, which was associated with a lower level of miR-155, indicating that the favourable effects of exercise on the circulating system may involve the participation of miR-155 in the molecular mechanism of improved vascular relaxation in muscular arteries
[31]. Despite an increase of miR-155 in blood and no relationship between miR-155 and VO
2max in healthy people after exercise training, the relationship between miR-155 and respiratory fitness in treatment-induced pulmonary patients remains to be determined
[12][32].
2.2.6. Others
A systematic review for breast cancer patients suggested that let-7, miR-20a, and miR-210 may serve as biomarkers for anthracycline-based cardiotoxicity during chemotherapy
[33]. In the rat model treated with doxorubicin at increasing doses, the level of cTnT was significantly elevated in the 18mg/kg dose. The decrease of let-7g levels accompanied it. Only a few studies investigated the change of let-7 in cardiotoxic patients after physical activity
[34]. By targeting
PTEN, miR-20a prevented exercise-associated CVD, but whether miR-20a played a role in reversing treatment-induced CVD remains unknown
[35]. Accumulating evidence suggested that the expression of miR-210 was inversely associated with VO
2max after physical activity. A possible explanation is that, during exercise, a hypoxic condition in cardiac muscle triggered angiogenesis through
HIF-1α-induced miR-210
[6]. Further efforts are required to establish a correlation between these miRNAs and physical and physiologic parameters in cancer survivors.
2.3. Cancer Cachexia
2.3.1. miR-21
Evidence suggested that miR-21 was upregulated in denervated muscles. miR-21, along with miR-206, was sufficient for the atrophy process by targeting
YY1 and
eIF4E3 [36]. He et al.
[37] showed that tumour-derived microvesicles containing miR-21 could induce apoptosis of muscle cells through Toll-like 7 receptors on murine myoblasts. Moreover, c-Jun N-terminal kinase activity is required to mediate this apoptotic response. This may shed light on how tumour cells promote loss of muscle mass. Taken together, miR-21 plays an important role in myogenesis and muscle mass gain. Other studies showed that low psoas muscle mass index was a prognostic biomarker for metastasis in colorectal cancer patients. The level of serum miR-21 was significantly elevated in colorectal cancer patients with low psoas muscle mass. This indicated that serum miR-21 quantification might help clinicians make decisions on developing intervention strategies for these patients
[38]. However, it should be noted that miR-21 also regulated muscle loss in a natural process, such as ageing
[39]. Additionally, evidence showed that levels of serum miR-21 and other miRNAs, such as miR-146a, miR-221, and miR-222, are unrelated to muscle bulk and fat reserve in athletes
[21]. Few studies were conducted to explore the relationship between miR-21 and muscle mass and strength gain induced by physical activity, which therefore require further investigation.
2.3.2. miR-378
In vivo study showed that training-induced change of miR-378 was positively associated with muscle mass gains. However, miR-378 was downregulated in low responders to resistance exercise amongst weightlifters. In contrast, an increase in miR-378 was not observed in high responders
[40]. Aside from the loss of muscle mass, cancer cachexia is also associated with a significant loss of adipose tissue, which is caused by enhanced lipolysis in adipocytes in some but not all patients. Kulyte et al.
[41] detected the expression of miR-378 in abdominal subcutaneous adipose tissue from gastrointestinal cancer patients and found that miR-378 was significantly elevated in cachectic cancer patients compared to non-cachectic patients. Overexpression of miR-378 was positively associated with loss of adipose tissue, which involved catecholamine-stimulated lipolysis in adipocytes. Inversely, inhibition of miR-378 can reduce the expression of
LIPE, PLIN1, and
PNPLA2 genes, which encoded key regulators for lipolysis and therefore decreased catecholamine-stimulated lipolysis. In vivo study showed that exercise-induced change in miR-378 abundance was positively correlated to muscle mass gain
[40].
2.3.3. miR-1 and miR-133
Although miR-1 and miR-133 are clustered on the same chromosomal loci, they have very distinct roles in transcriptional circuits and therefore differentially regulate skeletal muscle cell proliferation and differentiation. miR-1 promotes myogenesis by targeting transcriptional gene histone deacetylase 4 in muscle, while miR-133 enhances myoblast proliferation by inhibiting serum response factor
[42]. Moreover, miR-1 and miR-133 show different expression patterns in response to aerobic and resistance exercises. Both miRNAs were upregulated within three hours following an acute aerobic exercise bout but downregulated after four weeks of aerobic endurance training
[43]. Again they decreased after 6 h of resistance training and remained downregulated after a week of resistance exercise training
[43]. Recent work suggested that reduced IGF-1 protein was associated with upregulated miR-133 during the differentiation of C2C12 cells. Overexpression of miR-133 during differentiation significantly suppressed the expression of the IGF-1 receptor at the transcriptional level by downregulating the phosphorylation of
Akt. Moreover, the upregulation of miR-133 can be accelerated by the addition of IGF-1. These results indicated that metabolism-related factors played an important role in myogenesis
[43][44]. MiR-133a-deficient mice had a low maximal exercise capacity and low resting metabolic rate. The downregulated transcription of a set of mitochondrial biogenesis regulators, such as nuclear respiratory factor-1, transcription factor A, and so on, may explain lower mitochondrial mass and impaired exercise capacity in miR-133a-deficient mice. Six weeks of endurance training improved the expression of miR-133a and stimulated mitochondrial biogenesis in wild-type mice but failed to enhance the mitochondrial function in miR-133a-deficient mice
[45]. Mechanistic analysis showed IGF-1 receptor, the target of miR-133a, and the hyperactivation of
Akt signalling were involved in the lower transcription of the mitochondrial biogenesis regulators
[45]. These results suggested that miR-133a reduced exercise capacity by lowering mitochondrial function. However, few studies were conducted to establish the correlation between the expression of miR-1 and miR-133 and muscle mass or strength after exercise training.
2.4. Depression and Anxiety
2.4.1. let-7
Evidence showed that depressed patients with 3-month treatment with antidepressants reported an elevation of a series of let-7 family members in blood, suggesting let-7 may be a therapeutic target in depression
[46]. In a genetic rat model of depression, the decreased let-7 could increase the levels of the proinflammatory cytokine
IL-6, leading to depression. On the other hand, running can reduce the level of
IL-6 and rescue the expression of let-7i. This can elevate mood, thus exerting an antidepressant-like effect
[47]. Studies showed that the baseline level of let-7b in treatment-resistant depression patients was 40% lower than healthy controls. In comparison, let-7c was 50% lower in treatment-resistant depression patients who received electroconvulsive therapy when compared to healthy controls. Their role in depression may involve in
PI3k-Akt-mTOR signalling pathway
[48]. Hung et al.
[49] demonstrated that the level of let-7e before starting antidepressant treatment was inversely correlated to the severity of depression. The expression of let-7e was also altered by antidepressant treatment, but whether let-7e levels were affected by exercise training remains unknown.
2.4.2. miR-132 and miR-182
In vitro study showed that miR-132 and miR-182 reduced the expression of brain-derived neurotrophic factor (BDNF), which was greatly important in the aetiology of depression. Patients with depressive symptoms had lower BDNF levels and higher miR-132 and miR-182 levels in blood compared to healthy controls. This supports the role of miR-132 and miR-182 in regulating BDNF expression
[50]. Additionally, the self-rating depression scale (SDS) score was negatively correlated to serum BDNF levels but positively correlated to serum miR-132 levels. In contrast, the serum BDNF expression was reversely associated with the expression of miR-132/miR-182
[50]. In a rat model, downregulation of miR-132 can mitigate behavioural impairment of rats exposed to single prolonged stress, the mechanism of which may involve methyl GpG-binding protein 2, a positive mediator of BDNF regulated by miR-132-3p
[51]. These results indicated that miR-132 and miR-182 may play a critical role in the BDNF-mediated pathway and may serve as diagnostic or therapeutic targets of mental disorders. Although a change of miR-132 was observed after endurance training, which can also reduce depressive symptoms in humans, whether the positive effect of exercise on depression involved the miR-132 regulatory pathway remains to be determined.
2.4.3. miR-134
An enriched environment could ameliorate depressive-like behaviours caused by chronic unpredictable mild distress (CUMS). The
SIRT1/miR-134 signalling pathway regulates its downstream molecules, including synaptic plasticity proteins and BDNF expression in primary cultured hippocampal neurons. The protective effect of an enriched environment may be through activating the
SIRT1/miR-134 signalling pathway, remodelling the dendritic spine, altering synaptic ultrastructure, and increasing synaptic plasticity proteins and BDNF expression in the hippocampus
[52].
2.4.4. miR-34b/c
After exposure to NBI-27914, a specific corticotropin-releasing hormone receptor 1 (CRHR1) antagonist, which was correlated to trauma-induced anxiety, the rat model was observed for changes in the hypothalamic-pituitary adrenal (HPA) axis and anxiety-like behaviour. Bioinformatic analysis showed that CRHR1 was the target of miR-34b, and the overexpression of miR-34b negatively modulated CRHR1 mRNA in the primary hypothalamic neurons, mitigating the hyperactivity of the HPA axis and anxiety-like behaviour
[53]. Evidence also showed that miR-34b/c had a negative impact on the cognitive function of major depressive patients
[54]. However, few studies explored how physical exercise alleviates psychological symptoms by modulating the expression of miR-34b/c.
2.4.5. Others
In addition to miR-132 and miR-134, CUMS gives rise to depression-like behaviours via
SIRT1/miR-124, and swimming exercise can reverse the expression of SIRT1 protein and the expression of these miRNAs in CUMS mice. However, the correlation between depressive behaviour and epigenetic changes of hippocampal plasticity was not established
[55]. In major depressive disorder patients, intracellular miR-146a level was negatively correlated with the severity of depression; by contrast, miR-155 showed the opposite effect
[49]. However, their role in exercise-related prevention of depression requires further investigation.
3. The Challenge of miRNAs as Biomarkers in Exercise-based Cancer Rehabilitation
Albeit the promising prospect of miRNAs as potential biomarkers for exercise-based cancer rehabilitation, there are several issues remaining to be resolved: (1) clinical correlation between miRNAs expression in specific tissues and their expression in blood as well as physiological and physical parameters in cancer rehabilitation should be established due to the lack of related investigation; (2) the changing pattern of miRNAs differs from conventional biomarkers after exercise: NT-proBNP and hs-CRP increased after the marathon and stay elevated after 24h of race completion. By contrast, circulating miRNAs elevated instantly after the race and returned to prerace level or even lower 24h postrace. These results suggested that circulating release and clearance mechanisms of miRNAs may differ from traditional biomarkers and that miRNAs may serve as real-time and simultaneous instead of long-term markers of exercise-induced muscle adaptation
[56]. In addition, most studies investigated the effect of physical activity on miRNAs among healthy people or athletes, and few studies focus on the change of miRNAs on the deconditioned cancer patients during exercise. Therefore, it is necessary to conduct research exploring the effect of exercise-based rehabilitation on the dynamic change of miRNAs in cancer survivors; (3) miRNAs were associated with age, gender, training modality, or exercise regimen; we should, therefore, adjust these factors before we put miRNAs in use; (4) miRNAs at very low-level change are sufficient to cause an imbalance of human body by binding to multiple target genes
[14]. Thus, the target gene in a regulatory network needs to be fully elucidated. Thus, the target gene, as an auxiliary tool for miRNAs in rehabilitation, opens a new opportunity for future investigation.
This entry is adapted from the peer-reviewed paper 10.3390/life11121439