Benzo[a]pyrene (BaP) is a polycyclic aromatic hydrocarbon (PAH) and is used as an indicator of PAHs. It is primarily produced by burning of fossil fuels, wood and other organic materials.
The mechanism of epigenetic changes induced by BaP are mainly due to the formation of CpG-BPDE adducts, between metabolite of BaP—BPDE and CpG, which leads to changes in the level of 5-methylcytosine. BaP also acts through inhibition of DNA methyltransferases activity, as well as by increasing histone deacetylases HDACs, i.e., HDAC2 and HDAC3 activity. The aim of this entry is to discuss the mechanism of the epigenetic action of BaP on the basis of the latest publications.
- benzo(a)pyrene
- methylation level
- lung cancer
- smoke
- carcinogenicity
- histone modifications
1.Introduction
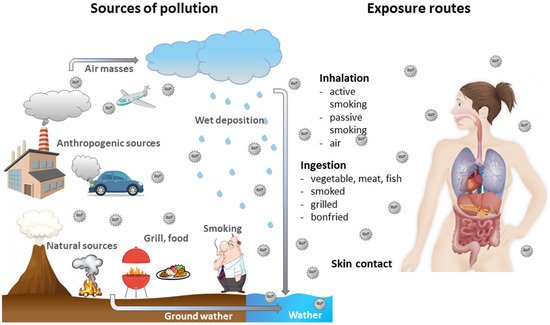
2. BaP Changes Global and Gene Specific DNA Methylation
Hypomethylation Induced by Benzo(a)pyrene and the Role of Poly(ADP-ribose) Glycohydrolase Silencing in DNA
3. The Intergenerational Toxic Effects on BaP Exposure Via Interference of the Circadian Rhythm
4. BaP Osteotoxicity and the Regulatory Roles of Epigenetic Factors; Intergenerational Osteotoxicity in Non-Exposed F3 Generation
5. Long-Term Exposure to BaP Inhibits Expression of ERα, CYP19a and VTG1 Genes and Is Toxic to Embryos and Sex Differentiation
6. Exposure to BaP in Mixture of PAHs Leads to Changes in DNA Modulation and RNA (hydroxy)methylation
7. Summary
| Type of Epigenetic Change | Observed Effects | References | |
|---|---|---|---|
| Global methylation | Increased global metylation: | Mouse embryonic fibroblast cells | [24] |
| Human bronchial epithelial cells (16HBE cells) | [20] | ||
| Mouse embryonic fibroblast cells | [24] | ||
| Normal human bronchial epithelial cells (NHBE) | [16] | ||
| Decreased global methylation: | Human bronchial epithelial cell line (16HBE) | [25] | |
| Zebrafish embryos | [23] | ||
| Zebrafish embryos | [26] | ||
| IRC mice | [27] | ||
| Children, whose mothers smoked during pregnancy | [36] | ||
| No changes in global methylation: | Human cells | [21] | |
| TK6 cells | [22] | ||
| Single gene promoters methylation and gene expression |
Increased methylation in gene promoter and decreased expression in gene promoter |
IFNγ—Jurkat cells and 53 women and childrenfrom the Columbia cohort | [37] |
| NR2B—C57BL mice | [29] | ||
| H19—302 reproductive-aged males (22–46 years old) | [38] | ||
| PER1—medaka fish Oryzias melastigma | [32] | ||
| Biotinidase and holocarboxylase synthetaseHuman Bronchial Epithelial Cells (16HBE) | [39] | ||
| 828 hypermethylated genes in rats | [28] | ||
| Decreased methylation in gene promoter |
3 227 hypomethylated genes in rats | [28] | |
| Decreased gene expression | ERα, CYP19a and VTG1 expression—Oryzias Melastigma | [34] | |
| Histone modifications | Reduction in acetylation levels on H3 and H4 histones |
Human bronchial epithelial cells (16HBE) | [39] |
| Increase in histone deacetylases HDAC2 and HDAC3 | Human bronchial epithelial cells (16HBE) | [39] | |
| Changes in H3K9 histone acetylation |
Breast cancer cells (MCF7) | [40] | |
| Changes in miRNA level |
Decreased expression | miR-506 in cancer cells (16HBE-T) | [41] |
| Increased expression | miR-638 in breast cancer (BRCA1) | [42] | |
| mikroRNA-29b, mikroRNA-26a-1* i mikroRNA-122* (HepG2 cells) | [43] | ||
This entry is adapted from the peer-reviewed paper 10.3390/ijms222413453
References
- IARC Working Groups Benzopyrene. Available online: https://monographs.iarc.who.int/wp-content/uploads/2018/06/mono100F-14.pdf (accessed on 14 December 2021).
- Susanto, A.D.; Yusril, N.; Zaini, J.; Nuwidya, F. Comparison of Serum Benzo(a)pyrene Diol Epoxide—Protein Adducts Level between Kretek Cigarette Smokers and Nonsmokers and the Related Factors. J. Nat. Sci. Biol. Med. 2021, 12, 52.
- Siddique, R.; Zahoor, A.F.; Ahmad, H.; Zahid, F.M.; Karrar, E. Impact of different cooking methods on polycyclic aromatic hydrocarbons in rabbit meat. Food Sci. Nutr. 2021, 9, 3219–3227.
- Migdał, W.; Walczycka, M.; Migdał, Ł. The Levels of Polycyclic Aromatic Hydrocarbons in Traditionally Smoked Cheeses in Poland. Polycycl. Aromat. Compd. 2020, 1–13.
- Schreiberová, M.; Vlasáková, L.; Vlček, O.; Šmejdířová, J.; Horálek, J.; Bieser, J. Benzopyrene in the Ambient Air in the Czech Republic: Emission Sources, Current and Long-Term Monitoring Analysis and Human Exposure. Atmosphere 2020, 11, 955.
- He, Y.; Yang, C.; He, W.; Xu, F. Nationwide health risk assessment of juvenile exposure to polycyclic aromatic hydrocarbons (PAHs) in the water body of Chinese lakes. Sci. Total Environ. 2020, 723, 138099.
- Cao, H.; Wang, C.; Liu, H.; Jia, W.; Sun, H. Enzyme activities during Benzopyrene degradation by the fungus Lasiodiplodia theobromae isolated from a polluted soil. Sci. Rep. 2020, 10, 865.
- Olgun, B.; Doğan, G. Polycyclic aromatic hydrocarbon concentrations in soils of greenhouses located in Aksu Antalya, Turkey. Water Sci. Technol. 2020, 81, 283–292.
- Amadou, A.; Praud, D.; Coudon, T.; Deygas, F.; Grassot, L.; Faure, E.; Couvidat, F.; Caudeville, J.; Bessagnet, B.; Salizzoni, P.; et al. Risk of breast cancer associated with long-term exposure to benzopyrene (BaP) air pollution: Evidence from the French E3N cohort study. Environ. Int. 2021, 149, 106399.
- Kim, K.E.; Cho, D.; Park, H.J. Air pollution and skin diseases: Adverse effects of airborne particulate matter on various skin diseases. Life Sci. 2016, 152, 126–134.
- Korsh, J.; Shen, A.; Aliano, K.; Davenport, T. Polycyclic Aromatic Hydrocarbons and Breast Cancer: A Review of the Literature. Breast Care 2015, 10, 316–318.
- Hamidi, E.N.; Hajeb, P.; Selamat, J.; Razis, A.F.A. Polycyclic Aromatic Hydrocarbons (PAHs) and their Bioaccessibility in Meat: A Tool for Assessing Human Cancer Risk. Asian Pac. J. Cancer Prev. 2016, 17, 15–23.
- Petit, P.; Maître, A.; Persoons, R.; Bicout, D.J. Lung cancer risk assessment for workers exposed to polycyclic aromatic hydrocarbons in various industries. Environ. Int. 2019, 124, 109–120.
- Birkett, N.; Al-Zoughool, M.; Bird, M.; Baan, R.A.; Zielinski, J.; Krewski, D. Overview of biological mechanisms of human carcinogens. J. Toxicol. Environ. Health Part B 2019, 22, 288–359.
- Martin, E.M.; Fry, R.C. Environmental Influences on the Epigenome: Exposure- Associated DNA Methylation in Human Populations. Annu. Rev. Public Health 2018, 39, 309–333.
- Hu, W.; Feng, Z.; Tang, M.-S. Preferential Carcinogen−DNA Adduct Formation at Codons 12 and 14 in the Human K-ras Gene and Their Possible Mechanisms. Biochemistry 2003, 42, 10012–10023.
- Wojciechowski, M.F.; Meehan, T. Inhibition of DNA methyltransferases in vitro by benzopyrene diol epoxide-modified substrates. J. Biol. Chem. 1984, 259, 9711–9716.
- Sadikovic, B.; Haines, T.R.; Butcher, D.T.; Rodenhiser, D.I. Chemically induced DNA hypomethylation in breast carcinoma cells detected by the amplification of intermethylated sites. Breast Cancer Res. 2004, 6, R329–R337.
- Sadikovic, B.; Rodenhiser, D.I. Benzopyrene exposure disrupts DNA methylation and growth dynamics in breast cancer cells. Toxicol. Appl. Pharmacol. 2006, 216, 458–468.
- Damiani, L.A.; Yingling, C.M.; Leng, S.; Romo, P.E.; Nakamura, J.; Belinsky, S.A. Carcinogen-Induced Gene Promoter Hypermethylation Is Mediated by DNMT1 and Causal for Transformation of Immortalized Bronchial Epithelial Cells. Cancer Res. 2008, 68, 9005–9014.
- Tommasi, S.; Kim, S.-I.; Zhong, X.; Wu, X.; Pfeifer, G.P.; Besaratinia, A. Investigating the Epigenetic Effects of a Prototype Smoke-Derived Carcinogen in Human Cells. PLoS ONE 2010, 5, e10594.
- Tabish, A.M.; Poels, K.; Hoet, P.; Godderis, L. Epigenetic Factors in Cancer Risk: Effect of Chemical Carcinogens on Global DNA Methylation Pattern in Human TK6 Cells. PLoS ONE 2012, 7, e34674.
- Corrales, J.; Fang, X.; Thornton, C.; Mei, W.; Barbazuk, W.; Duke, M.; Scheffler, B.; Willett, K. Effects on specific promoter DNA methylation in zebrafish embryos and larvae following benzopyrene exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 163, 37–46.
- Yauk, C.L.; Polyzos, A.; Rowan-Carroll, A.; Kortubash, I.; Williams, A.; Kovalchuk, O. Tandem repeat mutation, global DNA methylation, and regulation of DNA methyltransferases in cultured mouse embryonic fibroblast cells chronically exposed to chemicals with different modes of action. Environ. Mol. Mutagen. 2008, 49, 26–35.
- Huang, H.; Hu, G.; Cai, J.; Xia, B.; Liu, J.; Li, X.; Gao, W.; Zhang, J.; Liu, Y.; Zhuang, Z. Role of poly(ADP-ribose) glycohydrolase silencing in DNA hypomethylation induced by benzo(a)pyrene. Biochem. Biophys. Res. Commun. 2014, 452, 708–714.
- Fang, X.; Thornton, C.; Scheffler, B.E.; Willett, K.L. Benzopyrene decreases global and gene specific DNA methylation during zebrafish development. Environ. Toxicol. Pharmacol. 2013, 36, 40–50.
- Zhao, L.; Zhang, S.; An, X.; Tan, W.; Pang, D.; Ouyang, H. Toxicological effects of benzopyrene on DNA methylation of whole genome in ICR mice. Cell. Mol. Boil. 2015, 61, 115–119.
- Zhang, C.M.; Sun, Z.X.; Wang, Z.L.; Chen, J.S.; Chang, Z.; Zhu, L.; Ma, Z.H.; Peng, Y.J.; Xu, Z.A.; Wang, S.Q. Abnormal methylation of spermatozoa induced by benzo(a)pyrene in rats. Hum. Exp. Toxicol. 2019, 38, 846–856.
- Zhang, W.; Tian, F.; Zheng, J.; Li, S.; Qiang, M. Chronic Administration of Benzo(a)pyrene Induces Memory Impairment and Anxiety-Like Behavior and Increases of NR2B DNA Methylation. PLoS ONE 2016, 11, e0149574.
- Krishnaiah, S.Y.; Wu, G.; Altman, B.; Growe, J.; Rhoades, S.D.; Coldren, F.; Venkataraman, A.; Olarerin-George, A.O.; Francey, L.J.; Mukherjee, S.; et al. Clock Regulation of Metabolites Reveals Coupling between Transcription and Metabolism. Cell Metab. 2017, 25, 961–974.e4.
- Patke, A.; Murphy, P.J.; Onat, O.E.; Krieger, A.C.; Özçelik, T.; Campbell, S.S.; Young, M.W. Mutation of the Human Circadian Clock Gene CRY1 in Familial Delayed Sleep Phase Disorder. Cell 2017, 169, 203–215.e13.
- Yin, X.-H.; Liu, Y.; Zeb, R.; Chen, F.-Y.; Chen, H.-Y.; Wang, K.-J. The intergenerational toxic effects on offspring of medaka fish Oryzias melastigma from parental benzopyrene exposure via interference of the circadian rhythm. Environ. Pollut. 2020, 267, 115437.
- Mo, J.; Au, D.W.-T.; Guo, J.; Winkler, C.; Kong, R.Y.-C.; Seemann, F. Benzopyrene osteotoxicity and the regulatory roles of genetic and epigenetic factors: A review. Crit. Rev. Environ. Sci. Technol. 2021, 1–39.
- Sun, D.; Chen, Q.; Zhu, B.; Lan, Y.; Duan, S. Long-Term Exposure to BenzoPyrene Affects Sexual Differentiation and Embryos Toxicity in Three Generations of Marine Medaka (Oryzias Melastigma). Int. J. Environ. Res. Public Health 2020, 17, 970.
- Duca, R.-C.; Grova, N.; Ghosh, M.; Do, J.-M.; Hoet, P.H.M.; Vanoirbeek, J.A.J.; Appenzeller, B.M.R.; Godderis, L. Exposure to Polycyclic Aromatic Hydrocarbons Leads to Non-monotonic Modulation of DNA and RNA (hydroxy)methylation in a Rat Model. Sci. Rep. 2018, 8, 10577.
- Joubert, B.; Håberg, S.E.; Nilsen, R.M.; Wang, X.; Vollset, S.E.; Murphy, S.; Huang, Z.; Hoyo, C.; Midttun, Ø.; Uicab, L.C.; et al. 450K Epigenome-Wide Scan Identifies Differential DNA Methylation in Newborns Related to Maternal Smoking during Pregnancy. Environ. Health Perspect. 2012, 120, 1425–1431.
- Tang, W.-Y.; Levin, L.; Talaska, G.; Cheung, Y.Y.; Herbstman, J.; Tang, D.; Miller, R.L.; Perera, F.; Ho, S.-M. Maternal Exposure to Polycyclic Aromatic Hydrocarbons and 5’-CpG Methylation of Interferon-γ in Cord White Blood Cells. Environ. Health Perspect. 2012, 120, 1195–1200.
- Yang, J.; Lu, Z.; Liu, Z.; Wang, L.; Qiang, M. Methylation of Imprinted Genes in Sperm DNA Correlated to Urinary Polycyclic Aromatic Hydrocarbons (PAHs) Exposure Levels in Reproductive-Aged Men and the Birth Outcomes of the Offspring. Front. Genet. 2021, 11, 1706.
- Xia, B.; Yang, L.Q.; Huang, H.Y.; Pang, L.; Yang, X.F.; Yi, Y.J.; Ren, X.H.; Li, J.; Zhuang, Z.X.; Liu, J.J. Repression of Bio-tin-Related Proteins by BenzoPyrene-Induced Epigenetic Modifications in Human Bronchial Epithelial Cells. Int. J. Toxicol. 2016, 35, 336–343.
- Sadikovic, B.; Andrews, J.; Carter, D.; Robinson, J.; Rodenhiser, D.I. Genome-wide H3K9 Histone Acetylation Profiles Are Altered in Benzopyrene-treated MCF7 Breast Cancer Cells. J. Biol. Chem. 2008, 283, 4051–4060.
- Zhao, Y.; Liu, H.; Li, Y.; Wu, J.; Greenlee, A.R.; Yang, C.; Jiang, Y. The role of miR-506 in transformed 16HBE cells induced by anti-benzopyrene-trans-7,8-dihydrodiol-9,10-epoxide. Toxicol. Lett. 2011, 205, 320–326.
- Li, D.; Wang, Q.; Liu, C.; Duan, H.; Zeng, X.; Zhang, B.; Li, X.; Zhao, J.; Tang, S.; Li, Z.; et al. Aberrant Expression of miR-638 Contributes to Benzo(a)pyrene-Induced Human Cell Transformation. Toxicol. Sci. 2012, 125, 382–391.
- Lizárraga, D.; Gaj, S.; Brauers, K.J.; Timmermans, L.; Kleinjans, J.C.; Van Delft, J.H.M. Benzopyrene-Induced Changes in MicroRNA–mRNA Networks. Chem. Res. Toxicol. 2012, 25, 838–849.