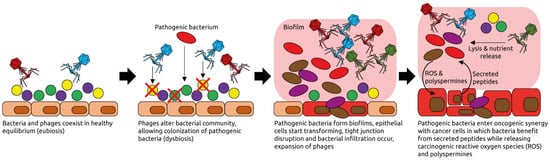
Viral infections as well as changes in the composition of the intestinal microbiota and virome have been linked to cancer. Moreover, the success of cancer immunotherapy with checkpoint inhibitors has been correlated with the intestinal microbial composition of patients. The transfer of feces—which contain mainly bacteria and their viruses (phages)—from immunotherapy responders to non-responders, known as fecal microbiota transplantation (FMT), has been shown to be able to convert some non-responders to responders. Since phages may also increase the response to immunotherapy, for example by inducing T cells cross-reacting with cancer antigens, modulating phage populations may provide a new avenue to improve immunotherapy responsiveness.
- virome
- microbiota
- cancer
- bacteriophages
- fecal microbiota transplantation
- checkpoint inhibitors
- immunotherapy
1. Introduction—The Human Virome
2. The Roles of the Eukaryotic Virome in Cancer
| Virus Family | Virus | Cancer Type | Observations |
|---|---|---|---|
| Papillomaviridae | HPV-16 | Anal | HPV, especially HPV-16, is a possible risk factor for anal and rectal cancer [8,9] and a significant prognostic marker, especially for locally advanced disease [10] |
| HPV | Bladder | HPV (different serotypes) may be linked to bladder cancer in a small number of cases [11] | |
| HPV-16, -18 | Cervical | Association between infection with high-risk HPV serotype (mainly HPV-16 and-18) and development of cervical cancer [11,12,13] | |
| HPV-18 | Colorectal | HPV, especially HPV-18, is a possible risk factor for colorectal cancer [14,15], however, another study found no association [16] | |
| HPV-16, -18, -26, -57 | Esophageal | HPV-16 is a risk factor for esophageal carcinoma [17,18]; HPV infection (mainly HPV-16, -18, -26 and-57) is common in esophageal carcinoma [19] | |
| HPV-16 | Head and neck (SCC) | HPV infection, especially HPV-16, is associated with head and neck cancer [11,20,21] and better long-term outcome [22] | |
| HPV-6 | Oral | Association of HPV-6 with oral cancer [23] | |
| HPV-16 | Prostate | Association of HPV-16 with prostate cancer [24] | |
| HPV-16, -18, -58 | Renal | Association of HPV-16, -18 and-58 with renal cell carcinoma [25] | |
| HPV-5, -8 | Skin and mucosal | Papillomavirus DNA frequently detected in skin-and mucosa-associated cancers [26]; HPV-5 and-8 are associated with epidermodysplasia verruciformis associated with a high risk of skin cancer [27,28] | |
| HPV-16 | Vulvar | Association between HPV, especially HPV-16, and vulvar squamous cell carcinoma [29] | |
| Herpesviridae | CMV (HHV5) | Colorectal | CMV DNA is more abundant cancer tissues compared to healthy tissues [30]; CMV-positive tumors in non-elderly patients are associated with increased disease-free survival rate [31]; specific genetic polymorphisms of CMV are linked to different clinical outcomes [32] |
| EBV (HHV4) | Colorectal | Possible association of EBV with colorectal carcinoma [33], however, no association found in another study [16] | |
| EBV (HHV4) | Esophageal | EBV is associated with esophageal cancer [34] | |
| EBV (HHV4) | Gastric | Possible involvement of EBV in gastric cancer and precursor lesions [35]; patients with EBV-positive gastric cancer had a better response to chemotherapy and better survival [36] | |
| EBV (HHV4) | Hepatic | EBV infections detected in HCC tissues [37] | |
| EBV (HHV4) | Lymphoma (Burkitt) | EBV infections contribute to Burkitt lymphoma [38] | |
| EBV (HHV4) | Lymphoma (DLBCL) | EBV RNA detected in B-cell lymphoma samples [39] | |
| EBV (HHV4) | Lymphoma (PTCL) | EBV expression associated with some subtypes of peripheral T-cell lymphomas [40] | |
| EBV (HHV4) | Oral | Higher proportion of EBV-positive oral squamous cell carcinoma in industrialized countries [41] | |
| EBV (HHV4) | Skin and mucosal | EBV DNA frequently detected in skin and mucosal cancers [26] | |
| HHV6 | Lymphoma (DLBCL) | HHV6 RNA detected in B-cell lymphoma samples [39] | |
| HHV6 | Malignant melanoma | HHV6 DNA frequently detected in malignant melanoma [26] | |
| HHV7 | Bladder | HHV7 DNA frequently detected in bladder cancer [26] | |
| HHV7 | Lymphoma (CTCL) | HHV7 DNA frequently detected in cutaneous T-cell lymphoma (Mycosis fungoides) [26] | |
| HHV7 | Oral | HHV7 DNA frequently detected in oral cavity cancer [26] | |
| HSV (HHV1/2) | Oral | Higher proportion of HSV-positive oral squamous cell carcinoma in industrialized countries [41] | |
| KSHV (HHV8) | Kaposi sarcoma | In HIV-infected individuals, KSHV infection is associated with Kaposi sarcoma [42] | |
| Polyomaviridae | BKV | Bladder | Possible association of BKV with bladder cancer [26] |
| BKV | Colorectal | Possible association of BKV with colorectal cancer [43,44], however, other studies found no association [16,45] | |
| JCV | Colorectal | JCV is associated with colorectal cancer [45,46] and may be involved in carcinogenesis [47], specifically in chromosomal instability [48]; JCV T-antigen is expressed in early-stage colorectal cancer [49], however, another study found no association [16] | |
| MCV | Merkel cell carcinoma | MCV is the major causative factor for Merkel cell carcinoma [50,51] | |
| Retroviridae | HIV | Anal | HIV-positive people have increased risk for anal cancer [52,53] and worse overall colostomy-free survival rates [54] |
| HIV | Cervical | Cervical cancer is more prevalent in HIV-positive individuals, likely because of increased susceptibility to HPV infection [55,56] | |
| HIV | Kaposi sarcoma | Kaposi sarcoma is more prevalent in HIV-positive individuals, likely because of increased susceptibility to KSHV infection [55,56] | |
| HIV | Lymphoma (NHL) | Aggressive B cell non-Hodgkin lymphoma is more prevalent in HIV-positive individuals, likely because of increased susceptibility to EBV infection [55,56] | |
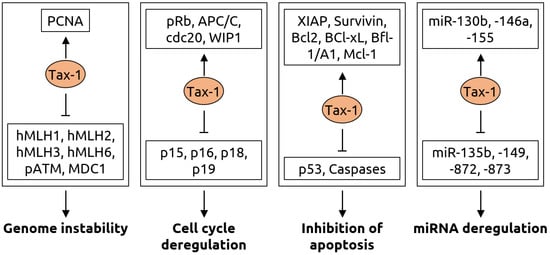
| HTLV-1 | Lymphoma (ATLL) | HTLV-1 induces adult T-cell leukemia/lymphoma in 5% of infected individuals [57] through random integration into the host genome [58] | |
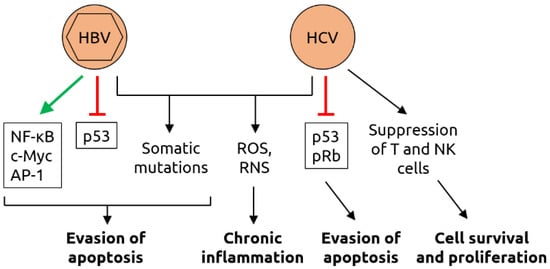
| Others | HBV | Bile duct | HBV is a risk factor for bile duct cancer [59] |
| HBV | Colorectal | Chronic HBV infection is a risk factor for colorectal cancer [60] | |
| HBV | Hepatic | Liver cancer is associated with HBV [11] | |
| HBV | Pancreatic | Chronic HBV infection [61] or past exposure [62] are risk factors for pancreatic cancer | |
| HCV | Bile duct | HCV is a risk factor for bile duct cancer [59] | |
| HCV | Hepatic | Liver cancer is associated with HCV [11] | |
| TTV | Hepatic | TTV is a risk factor for hepatocellular carcinoma [63] | |
| HBoV | Colorectal | Some colorectal cancers are associated with HBoV [64] | |
| HBoV | Lung | Some lung cancers are associated with HBoV [64] | |
| HBoV | Tonsillar | Association of HBoV with tonsil squamous cell carcinoma [65] | |
| Orthobunyaviruses | Colorectal | High abundance of orthobunyaviruses in colorectal cancer [66] | |
| Parvoviruses | Skin | Parvovirus DNA frequently detected in skin-associated cancers [26] | |
| Anelloviruses | Mucosal | Anellovirus DNA frequently detected in mucosal cancers [26] | |
| Anelloviruses | Leukemias | Anellovirus DNA frequently detected in leukemias [26] |
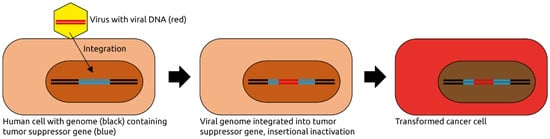
2.1. Papillomaviridae
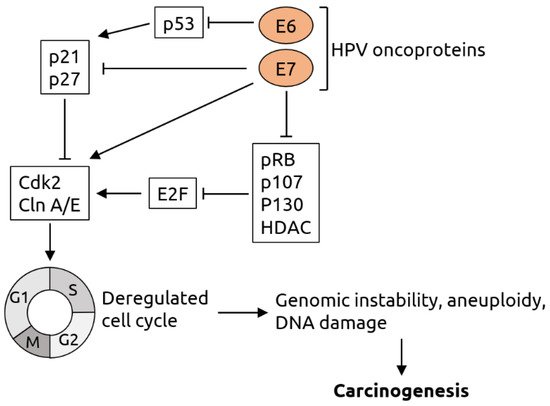
2.2. Herpesviridae
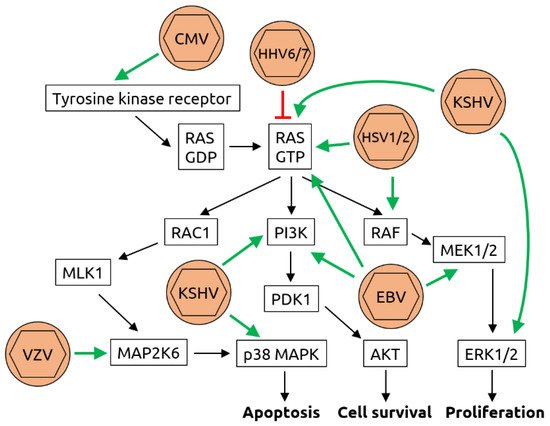
2.3. Polyomaviridae
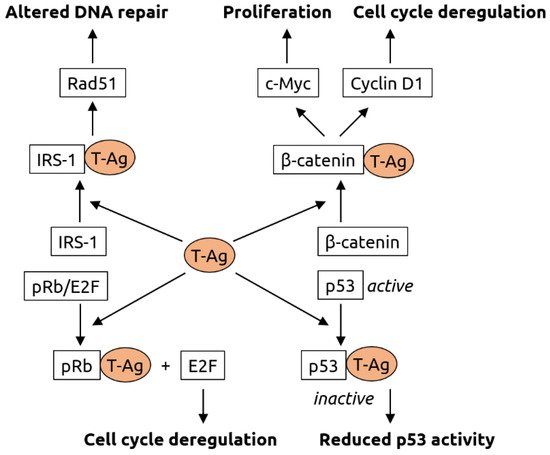
2.4. Retroviridae
2.5. Other Viruses
3. The Human Intestinal Virome and Its Links to Cancer
This entry is adapted from the peer-reviewed paper 10.3390/microorganisms9122538