Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Obstetrics & Gynaecology
|
Oncology
Female oncofertility is recently implemented conception in the female fertility treatment. It includes the therapeutic approaches to preserve the fertility competence in cancer patients.
- chemotherapy
- fertility preservation
- gonadotoxic
- oncofertility
- oocyte quality
1. Introduction
In the past two decades, substantial advances in early diagnosis and cancer treatment have resulted in an approximately 80% 5-year survival rate in young oncological patients [1,2], leading to a rise in number of female childhood cancer survivors (CCS) [3]. However, oncologic treatment usually requires extensive chemotherapy and/or radiotherapy, which are indicated to be distinctively ovotoxic, resulting in premature ovarian insufficiency (POI) and consequent infertility [4,5,6,7]. Approximately, 30% of children who were exposed to chemo- and/or radio-therapy develop gonadal dysfunction [8]. The incidence of POI in CCS is estimated as high as 8–10% [4].
Although the mechanism is not fully elucidated yet, current data demonstrate that chemotherapeutic agents, especially alkylating ones, interfere with DNA replication and cell division [9], massively activate the primordial follicles (PFs) [10,11], cause stroma atresia [12], and damage the vascularity in ovaries [13]. The radiation is also harmful to oocytes as its low dose of less than 2 Gy can destroy 50% of primordial follicles [7,14].
This fertility-compromised status has been well-documented to cause emotional distress and poor quality of life [15,16,17,18]. It was reported that there will be approximately 100 million women at risk of chemotherapy-induced ovarian impairment in 2025 [19]. In this context, preserving the fertility and quality of life of CCS has received considerable concerns. During the last two decades, FP with several effective approaches has been significantly developed, and represents a beneficial option to help hundreds of oncological women have genetic offspring. Furthermore, increasing studies are attempting to clarify the mechanisms and outcomes of chemo- and radio-therapy impacts on the ovarian reserve and oocyte quality, to develop protective methods as well as to improve therapeutic approaches in FP.
2. Impact of Chemo- and Radio-Therapy on Follicle Quantity
2.1. Clinical Data Describing the Impact of Chemo- and Radio-Therapy on Ovarian Function
A growing number of studies have demonstrated that the pregnancy rate and live birth rate in female CCS are lower compared to those of their siblings and general population controls. The results of these studies are summarized in Table 1.
Table 1. Summary of published clinical studies describing chemo- and radio-therapy on ovarian function.
| Authors | Number of CCS | Age | Exposure Agent a | Radiation | Effects | |
|---|---|---|---|---|---|---|
| Clinical | Laboratory Test | |||||
| Berjeb et al. (2021) [20] | 66 | 15–40 (26.7 ± 6.8) |
Bleomycin, etoposide, doxorubicin, cyclophosphamide, vincristine, procarbazine, doxorubicin, vinblastine, dacarbazine | No | N/A | ↓ AMH |
| Filippi et al. (2021) [21] | 90 | 21.3 ± 5.4 | Bleomycin, cisplatin, bleomycin, dacarbazine-vinblastine | Yes/No b | ↑ POI rate (21% of treated women) | |
| Gini et al. (2019) [22] | 97 | 16–50 (median: 28) |
Doxorubicin, cyclophosphamide, vincristine, bleomycin | Yes | ↑ Amenorrhea | N/A |
| Lehmann et al. (2019) [23] | 444 | ≤40 | N/A | Yes/No | N/A | ↑ LH ↑ FSH ↓ E2 |
| Anderson et al. (2018) [4] | 23,201 | ≤39 | N/A | N/A | ↓ Pregnancy rate (↓ 38%) | |
| Shandley et al. (2018) [24] | 1090 | 20–35 (median: 26) |
N/A | No | N/A | ↓ AFC↓ AMH |
| Sinha et al. (2018) [25] | 88 | 24–43 (median: 35) |
Taxotere, cyclophosphamide, carboplatin, fluorouracil, epirubicin | No | N/A | ↓ AFC |
| Al-Rawi et al. (2018) [26] | 58 | 25–45 (38.83 ± 4.74) |
Anthracycline, cyclophosphamide | No | N/A | ↓ AFC↓ E2 ↑ LH |
| Aderson et al. (2018) [27] | 67 | 18–45 | Doxorubicin, bleomycin, vinblastine, and dacarbazine | No | N/A | ↓ AMH ↑ FSH ↓ E2 |
| Levine et al. (2018) [28] | 2930 | 18–58 (median: 32) |
Alkylating agent, procarbazine | Yes/No | ↑ POI rate (9.1% of treated women) | N/A |
| Armuand et al. (2017) [29] | 552 | ≥13 | N/A | N/A | ↓ The probability of having a first live birth | N/A |
| Chemaitilly et al. (2017) [30] | 988 | 18–45 (median: 31.7) |
Alkylating agents | Yes | ↑ POI rate (10.9% of treated women) | N/A |
| D’Avila et al. (2017) [31] | 52 | 27–40 (35.3 ± 3.8) |
Cyclophosphamide | No | ↑ Amenorrhea | ↓ AFC↓ AMH ↑ FSH |
| Abir et al. (2016) [32] | 20 | 5–18 | Alkylating agents, bleomycin, cisplatin, vincristine, etoposide, carboplatin, doxorubicin, etopside, doxorubicin, bleomycin, vinblastine, dacarbazine. | No | ↑ Atretic follicles↓ Oocyte maturation | N/A |
| Hamy et al. (2016) [33] | 134 | 26–43 (median: 36) |
Anthracyclines, taxane | No | N/A | ↓ AMH |
| Even-Or et al. (2016) [34] | 35 | 13–36 (median: 25.5) |
Melphalan | No | N/A | ↓ AMH ↑ FSH ↓ LH |
| Gupta et al. (2016) [35] | 16 | 11–18 (median: 14.3) |
Doxorubicin, cyclophosphamide, cisplatin | No | ↑ Amenorrhea | ↓ AMH |
| Chow et al. (2016) [5] | 5298 | 15–44 | Busulfan, carboplatin, carmustine, chlorambucil, chlormethine, cisplatin, cyclophosphamide, dacarbazine, ifosfamide, lomustine, melphalan, procarbazine, temozolomide | Yes/No | ↓ Pregnancy rate | N/A |
| Thomas-Teinturier et al. (2015) [36] | 105 | 18–39 (median: 21.5) |
Cyclophosphamide, ifosfamide | Yes | N/A | ↓AMH ↑ FSH |
| Behringer et al. (2012) [37] | 106 | 18–39 (28 ± 7) |
Bleomycin, etoposide, doxorubicin, cyclophosphamide, vincristine, procarbazine, doxorubicin, bleomycin, vinblastine, dacarbazine | N/A | N/A | ↓ AMH ↑ FSH |
| Green et al. (2009) [38] | 5149 | 15–44 | Alkylating agents | Yes/No | ↓ Pregnancy rate | N/A |
a: All chemotherapeutic agents exposed that all included patients were exposed to are listed in each study. b: Some patients treated by both radiation and chemotherapy, but some patients were treated only with chemotherapy. ↓: Decreased. ↑: Increased. AFC: antral follicle count, AMH: anti-Müllerian hormone, E2: estradiol, FSH: follicle-stimulating hormone, LH: luteinizing hormone, N/A: not available or not applicable, POI: premature ovarian insufficiency.
In a longitudinal study including 66 patients undergoing chemotherapy, the AMH levels were decreased significantly (0.90 ± 1.55 compared to 2.61 ± 2.20 ng/mL before treatment) after chemotherapy using BEACOPP protocol (Bleomycin, Etoposide, Doxorubicin, Cyclophosphamide, Vincristine, Procarbazine, and Prednisolone) during a following period of 16.8 ± 9.3 months. In the ABVD protocol (Doxorubicin, Bleomycin, Vinblastine, and Dacarbazine), the AMH levels prior and after treatment were not statistically different (4.38 ± 3.39 vs. 4.27 ± 3.09 ng/mL, p = 0.753) [20]. Another study recorded the rates of diminished ovarian reserve and POI after chemotherapy as 39% (35/90) and 21% (19/90), respectively [21].
According to a population-based analysis, the overall likelihood of pregnancy in female CCSs aged under 40 is a 38% lower than that in the general population of women [4]. In another study, laboratory results show impairment in the concentration of female gonadal-related hormones (LH, FSH, and estradiol) in 24.3% (97/444) of female CCSs who were younger than 40 years of age [23]. According to a cohort study of 552 female CCSs in Sweden, the hazard ratio (HR) for having a first live birth in CCSs with malignancy of the eye, central nervous system tumors, and leukemia, is statistically lower than in healthy controls [29]. In 2930 CCSs, 110 survivors encountered POI with the value of 10.3 as an odds ratio compared to their healthy siblings, resulting in lower birth rates in their thirties [28]. In another report, the relative likelihood of 5149 CCSs achieving pregnancy is 0.81 (95% CI, 0.73 to 0.90; p < 0.001) compared with that of female siblings [38]. In a large sample cohort study including 5298 female five-year cancer survivors, their likelihood of having a pregnancy is significantly lower than their siblings (HR 0.85, 95%: 0.74–0.98; p = 0.023) [5]. The effects of the alkylating drugs and cisplatin on ovarian functions show a dose-dependent manner [5]. In a systematic review including 5607 female CCSs, the prevalence of amenorrhea ranges from 0% to 83% [6]. Exposure to alkylating agents and older age at treatment are detected as the decisive factors contributing to ovarian dysfunction [6].
2.2. Mechanism of Chemo- and Radio-Therapy Induction of Follicular Loss
To develop new therapies of FP and fertoprotective agents, numerous studies have described the possible mechanisms in which chemo- and radio-therapy induce ovarian damage. As typical chemotherapy protocols often consist of several agents, determining the ovarian impairment caused by each type of antitumor drug in clinical studies is challenging. Consequently, the conceptual effect of a single drug on the ovary is usually clarified by in vitro cell culture, ovarian tissue culture, or in vivo animal models, and human ovarian tissue culture followed by xenotransplantation [39,40].
Chemotherapeutic agents are generally divided into five categories: alkylating agents (cyclophosphamide, procarbazine, and busulfan), platinum-based compounds (cisplatin and carboplatin), anthracycline antibiotics (doxorubicin and bleomycin), antimetabolites (methotrexate and 5-fluorouracil), and vinca alkaloids (vincristine and vinblastine). The first three groups are demonstrated to damage ovaries by inducing DNA alterations, leading to follicular apoptosis [41,42]. Among these, alkylating agents are supposed to be most ovotoxic, causing significant follicular loss [6,43]. The last two groups are indicated to have a low risk to ovarian function [42]. However, some data show that vinca alkaloids, due to their suppression of microtubule dynamics, induce a vascular impairment, leading to ovarian dysfunction [44,45]. Three major mechanisms were proposed by several scientific groups (Figure 1).
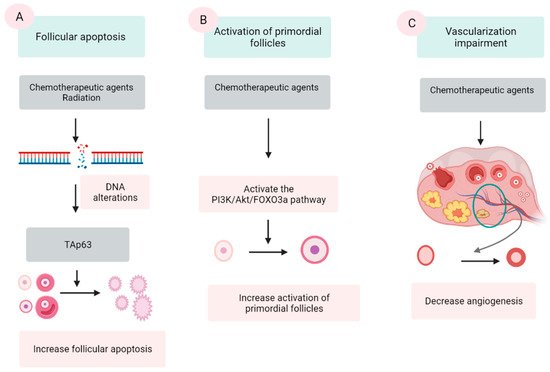
Figure 1. Three mechanisms of chemo- and radio-therapy-induced follicular quantity depletion: enhancement of apoptosis, accelerated activation of PFs. (A) DNA alterations induced by chemotherapeutic agents and radiation activates TAp53 protein, leading to the apoptosis. (B) Chemotherapeutic agents activate the phosphoinositide 3-kinase (PI3K)/Akt/forkhead box protein O3a (FOXO3a), which in turn induce the activation of PFs, resulting in the extensive loss of PFs. (C) Chemotherapeutic agents impair the epithelial tissue of vessels in the ovary, resulting in a reduction in the vascularization.
2.2.1. Follicular Apoptosis after Chemotherapy
The extensive apoptosis of ovarian follicles, especially PFs, after DNA alterations and/or oxidative stress is the most commonly described mechanism in chemotherapy-induced ovarian failure [41,46,47]. Several agents in the antitumor protocols, especially alkylating agents, are demonstrated to cause DNA lesions in both oocyte and granulosa cells (GCs). Among these lesions, double-stranded DNA breaks are among the most severe [39,42]. The accumulation of DNA strand breaks that could not be repaired by the DNA repairing system induces the apoptotic intracellular pathways, resulting in cellular apoptosis [42]. p63 protein (TAp63 isoform), Bcl2-associated X (BAX) protein and the BCL-2 antagonist killer (BAK) protein activator, is the major protein that mediate this mechanism [48,49].
Culture with cyclophosphamide [46] as well as in vivo cyclophosphamide injection [50] of mice’s ovaries induces DNA damage and subsequent follicle apoptosis. Cisplatin also causes DNA impairment and PFs’ apoptosis in both newborn and adult mouse ovaries [41]. Cyclophosphamide treatment substantially decreases the number of PFs, primary follicles, and secondary follicles with an elevated number of atretic follicles compared with control animals [13]. In another experiment, intraperitoneal injection of cyclophosphamide and cisplatin caused a significantly destructive effect on the PFs pool [51]. However, in mice with gene deletion of PUMA, a member of BCL-2 protein family, the PFs are retained after the treatment by both cyclophosphamide and cisplatin [51]. In a human ovarian xenograft model, cyclophosphamide [52], cisplatin [53], and doxorubicin [39] elevated DNA double-stranded breaks and resulted in a significant follicle loss.
The effect of antitumoral drugs on ovarian function is the follicle-specific magnitude and is associated with the category of the drugs [12]. Some studies have declared that apoptosis occurred only in GCs of growing follicles, but not in PFs by TUNEL staining after treatment of cyclophosphamide or cisplatin [54,55]. Other results insist that TUNEL and/or γH2AX staining are positive in oocytes but not in the GCs of PFs [46,50,56]. In another experiment, culturing ovaries with cisplatin or carboplatin decreases the number of follicles of all stages, but the most obvious reduction is observed in PFs [57]. One study reported that culturing of neonatal mice ovaries in cisplatin or doxorubicin significantly decreased the number of follicles at all stages [58]. However, the apoptosis evidence in the TUNEL analysis is not positive in the PFs, only in the growing follicles [58].
2.2.2. Activation of PFs Induced by Chemotherapy
An additional suggested mechanism for ovarian impairment after oncological treatment is the accelerated activation of PFs. Several scientific groups have confirmed that chemotherapy causes massive activation of PFs in affected ovaries via a phosphoinositide 3-kinase/protein kinase B/forkhead box protein O3a (PI3K/AKT/FOXO3a) pathway [54,59,60,61].
Mice administered intraperitoneally with cisplatin show a substantially decreased number of PFs along with higher numbers of early growing follicles and the signal of the key proteins in the PTEN/Akt/FOXO3a [59]. Other studies also revealed increased phosphorylation of Akt, mTORC, and downstream proteins followed by PF reduction in cyclophosphamide-treated mice [54,62]. In mice, doxorubicin causes detrimental effects on ovaries through both atresia and overactivation in PFs [63]. The same effects are found in mice treated with cisplatin [59]. In another experiment using neonatal mouse ovaries cultured with cisplatin or doxorubicin, PFs decrease without the evidence of apoptosis in TUNEL analysis, suggesting the etiology of PF reduction by overactivation [58]. In terms of human ovarian follicles, exposure to cyclophosphamide metabolites in vitro also induces PFs’ activation [61]. Furthermore, a cohort study of 96 female CCSs who were treated with alkylating agent revealed PFs activation in vivo and a remarkably suppressed nuclear expression of FOXO3a occurring in ovaries of these patients [64].
In consistence with this hypothesis, many experiments have indicated that inhibiting the PI3K pathway by several agents including rapamycin, ammonium trichloro (dioxoethylene-o,o′) tellurate (AS101), anti-Müllerian hormone (AMH), and melatonin, could prevent PF’s activation after chemotherapy [54,55,60,65,66,67,68].
Although this mechanism has been widely accepted, recent literature has raised the argument that activating PFs might not be the major or a specific cause of chemotherapy-induced PF loss [11]. Accordingly, the authors doubt that a growing follicles to PFs ratio calculation were not the correct parameter for a sign of PFs’ activation, because elimination of PFs could occur due to a deleterious effect. In an experiment, after culture of intact mouse ovaries with the metabolite agent cyclophosphamide, the number of PFs decreased along with increased levels of apoptotic markers BAX and cPARP. Meanwhile, there was no significant change in the number of primary follicles. In combination with the TUNEL staining’s results, this study indicated that the decrease in PFs was not due to their activation but the apoptosis in PFs [46]. A recent study demonstrated the depletion of PFs after cyclophosphamide exposure in a human ovarian xenograft model, utilizing triggering of proapoptotic pathways without evidence of PFs activation, and indicated that apoptosis was the main mechanism of PFs’ depletion [69].
2.2.3. Vascularization Impairment
Another proposed mechanism is the alteration in angiogenesis and stroma supporting the gonadal cells after exposure to chemotherapeutic agents [70,71]. Cyclophosphamide treatment shows induction of inflammation and enhanced expression of stromal cell-derived factor 1 (SDF-1), a factor related to follicular atresia, which presents in the granulosa, theca cells, and luteinized cells [72]. In human ovaries, histological analyses of ovaries from cancer survivors show the presence of damaged cortical blood vessels and proliferation of small vessels (neovascularization). Furthermore, the muscular layer in blood vessels becomes thicker, leading to limited blood circulation. The cortex presents fibrotic focal areas along with disappearance of follicles [73]. During in vivo monitoring, an evident reduction in ovarian circulation and spasm of small vessels are noted after the administration of doxorubicin [74]. In vitro human ovarian tissue culture with doxorubicin followed by xenograft to immunodeficient mice has a lower vascular density and higher microvascular compromise compared with controls [39]. In addition, one study assessing human ovarian tissue shows that both alkylating and nonalkylating drugs affect ovarian stromal function, leading to a substantial decrease in estradiol production [75].
2.2.4. Radiation
Regarding radiotherapy, the human oocyte is very sensitive to radiation, and a dose as low and less than 2 Gy for pelvic radiation can destroy 50% of PFs [76]. The position of radiation is one determinant factor of the degree of ovarian damage. The rate of POI in patients who experienced total body radiation and pelvic irradiation are 90% and 97%, respectively [77]. In addition, factors such as patient age and radiation dose are also important contributors [73,77]. Aging patients are more vulnerable to radiation compared with younger girls, due to the age-related decline in the follicle population [78]. The dose causing ovarian dysfunction in children is 1–2 Gy, whereas in adults it is as low as 0.4–0.6 Gy [79]. Based on an analysis from five centers conducting ovarian tissue cryopreservation (OTC), the live birth rates after OTC in patients undergoing pelvic irradiation reduced significantly in a dose-dependent manner [80].
The proposed mechanism of follicle depletion is the radiation-provoked ionizing damage of DNA [81]. This alteration also activates TAp63 protein, leading to destruction of PFs [49]. In terms of late effects, vasculature damage and stromal fibrosis following tissue hypoxia could be another mechanism [77]. This can result in ovarian atrophy and subsequent tissue dysfunction [73].
This entry is adapted from the peer-reviewed paper 10.3390/jcm10235690
This entry is offline, you can click here to edit this entry!