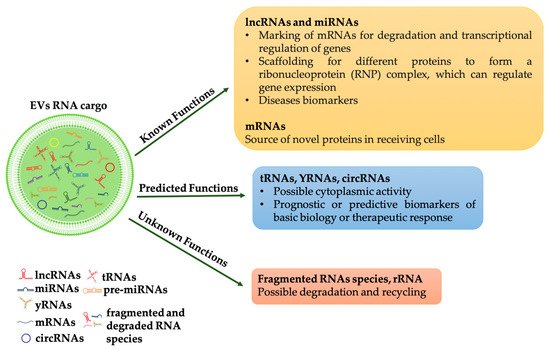
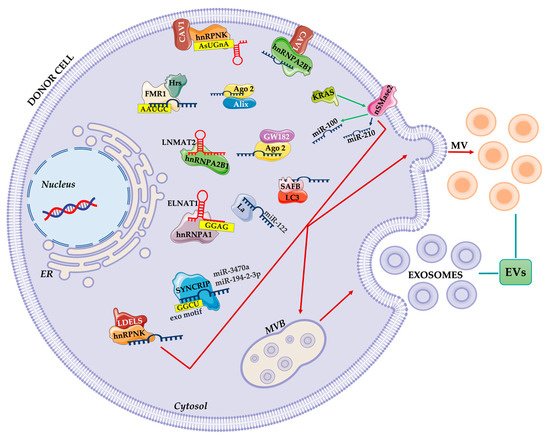
2. The Effects of the Horizontal Transfer of EVs Derived RNAs
Many studies demonstrated the functional role of the RNA-cargo that, selectively packaged inside EVs, are transferred to target cells. Firstly, the transfer of exosomal mRNAs and miRNAs was reported by Valadi et al. in 2007, which revealed a “novel mechanism of genetic exchange between cells” [
26]. Since that study, numerous papers have demonstrated that the horizontal transfer of exosomes cargo can modulate target cells behaviors. Studies on tumor cells revealed as exosome-mediated RNA transfer may control tumor growth, affect its microenvironment, and promote metastases.
It is well known that a central event in tumor progression is the induction of angiogenesis. Focusing on this pathway it has been demonstrated that glioblastoma, chronic myelogenous leukemia, and breast cancer derived EVs can reprogram endothelial cells through horizontal transfer of their miRNAs cargo [
27,
28]. The involvement of cancer derived EVs in facilitating brain infiltration is worth mentioning. Lu and collaborators have recently demonstrated that exosomes derived by highly brain metastatic breast cancer cells are able to destroy the blood–brain barrier through its lncRNA
GS1-600G8.5 [
29,
30]. Exosomal miRNAs have a role also in tumor drug resistance. Mao et al. support this hypothesis demonstrating that Adriamycin-resistant breast cancer cells deliver specific miRNAs through exosomes thus promoting drug resistance in neighboring cells [
31]. Quin and collaborators demonstrated that exosomes derived from cisplatin-resistant lung cancer cell line A549 induce drug resistance in receiving cells. MiRNA profile identified the
miR100-5p as the mediator of this process [
32].
In addition, exosome-transported lncRNAs may participate in drug resistance induction. A recent paper by Wang et al. proved that the exosome-mediated transfer of lncRNA
H19 induces doxorubicin resistance in breast cancer [
33]. The authors showed that drug-resistant cells release exosomes enriched in
lncH19; these exosomes increase the chemoresistance of doxorubicin once internalized by sensitive cells. Moreover, downregulation of
H19 in sensitive cells ablated this effect thus confirming the direct role of the long non-coding RNA [
33]. EVs released into the tumor microenvironment strongly affect metastatic niche. For example, the prometastatic miRNA
miR-9 and
miR-155, carried respectively by breast cancer derived exosomes and pancreatic cancer derived microvesicles, are able to reprogram fibroblast to cancer associated fibroblast (CAF) phenotype thus promoting tumor progression [
34,
35]. While the EV-mediated delivery of the
miR-105 and
miR-122 reprogram CAFs metabolism to sustain tumor growth [
36,
37]. Moreover, cancer derived EVs can reprogram immune cells thus preventing immunosurveillance and promoting immunotolerance in cancer microenvironment, as revised by Graner [
38]. In addition, EVs derived RNAs profiling can be useful as a prognostic indicator to therapeutic response. For example,
miR196a-5p and
miR-501-3p were significantly downregulated in exosomes isolated from the urine of prostate cancer patients [
39], while
let7-b and
miR-18a, isolated from plasma of multiple myeloma patients, were associated with overall survival [
40]. As expected, the effects of the EV-transported RNA are not limited to the tumor context.
Barile and colleagues revealed the cardioprotective role of the different miRNAs transported by EVs derived from cardiac progenitor cells (CPC). They demonstrated that
miR-210 inhibits cardiomyocyte apoptosis by targeting Ephrin A3 (cell surface GPI-bound ligand for Eph receptors) and PTP1b (protein-tyrosine phosphatase 1b). Moreover, EVs derived
miR132 stimulates angiogenesis acting on RasGAP-p120, Ras GTPase activating protein p120 [
41]. Furthermore, CPC derived exosomal
miR21 exert similar effects preventing cell apoptosis by targeting PDCD4 (Programmed Cell Death 4) [
42]. Similarly, Gray and collaborators, through microarray analysis of exosomes derived from hypoxic CPC, identified 11 miRNAs that improve cardiac function stimulating tube formation of endothelial cells and reducing fibrosis [
43].
Regarding differentiation, it has been demonstrated that exosomes released from Neural stem/progenitor cells (NPCs) have an important role in neurogenesis; Ma and collaborators demonstrated that mouse cortical NPCs, isolated from fetal brain, promote neuronal differentiation through exosomal
miR-21a [
44].
Adipose tissue is an excellent resource for circulating exosomal miRNAs that have a role in regulating liver gene expression, as well as affecting obesity or diabetes. Adipose-derived exosomal
miR-99b has been demonstrated to control in vivo fibroblast growth factor 21 (FGF21) production [
45]. While exosomes secreted by adipose tissue macrophages transfer miRNAs modulating, in vivo and in vitro, insulin sensitivity and glucose homeostasis [
46].
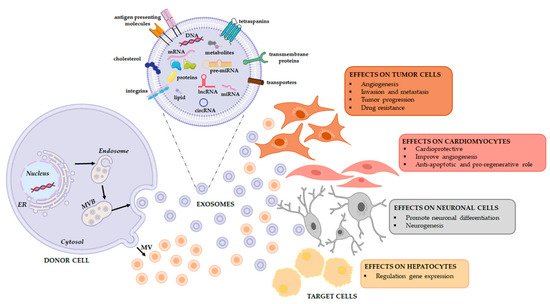
All these experiments suggest that exosomes may be a vehicle of therapeutic non-coding RNAs in physiological and pathological conditions as well as in the field of regenerative therapy (Figure 2). To this end, a comprehensive analysis of the RNA loading mechanisms is appropriate.
Figure 2. Extracellular vesicles release and their functional effects on target cells. EVs are a heterogeneous population, both in form and content since their cargo is strictly dependent on the pathophysiological conditions of the cell at the exact moment in which it produces the vesicle. When studying the complexity of the EV-mediated cell-cell communication, it is necessary to evaluate that the same vesicle, i.e., the same message, can be interpreted differently depending on the cytotype that receives it. This will depend in good part on the gene expression profile of the recipient cell.