1. Introduction
The tumor and the immune cell interaction are critical areas of study in cancer growth and progression. Macrophages have a controversial role in the tumor microenvironment with both anti- and pro- tumoral effects [
1]. The gaseous transmitter and signaling molecule nitric oxide (NO) and the expression of inducible nitric oxide synthase (iNOS) that produces NO are noted to have dual roles in cancer, which is to either promote or inhibit tumor growth [
2,
3]. Therefore, NO shows potential either as a therapeutic agent in its own right or as a target molecule in cancer therapies.
NO is a small signaling molecule that is synthesized by three NO synthases (NOS) isoforms. The two isoforms neuronal NOS (nNOS or NOS I), and endothelial NOS (eNOS or NOS III) are constitutive. The third NOS isoform (NOS 2 or inducible NOS, iNOS) is negligible in resting cells and is induced by cytokines and bacterial lipopolysaccharide (LPS) [
3,
4]. NO plays an important role in cell growth, differentiation, and apoptosis [
3,
5]. The enzyme iNOS produces NO through the conversion of L-arginine into citrulline utilizing NADPH and oxygen. NO may also be associated with resistance to apoptosis [
6] and immune escape [
7]. iNOS is induced by inflammatory cytokines [
8] and is transcriptionally regulated [
9]. It is well established and acknowledged that NO’s role in cancer depends on its concentration, exposure duration in cells, cell-specific sensitivities, iNOS localization in tissues, and extracellular conditions [
10,
11,
12].
Antitumor effects of NO have been demonstrated even though its pro-tumor effects have largely dominated the scenarios. In terms of the tumorigenic effects, NO contributes to tumor growth and metastasis, regulates metabolism by the Warburg effect, and promotes cancer growth via high glycolytic activity [
13]. Furthermore, antitumor iNOS activity is related to its cytotoxicity and immunogenic effects [
14]. Therefore, NO-releasing hybrids are subjects of intensive investigations as potential anticancer drugs, either as single cytotoxic agents or in combination with standard radio- and chemotherapy [
13].
More recently, studies relate the chemo- and immunoresistance [
7] in cancer cells with NO as a mediator for the events in the tumor microenvironment (TME) and as a bonafide molecular target [
15,
16].
2. Therapeutic Approaches Utilizing Macrophage-Derived iNOS/NO in Cancer
Antitumor strategies targeting TAMs include potential mechanisms of lowering TAM survival, reducing macrophage recruitment, and switching M2-like TAMs into an M1-like phenotype [
51]. Utilizing the iNOS derived-NO or exogenous NO delivery, there has been some success with nano-therapeutics. In the tumor microenvironment, the nanoparticles of self-assembled poly(L-arginine) are taken up by the activated macrophages, followed by the hydrolytic release of L-arginine, and conversion to NO by the iNOS of the TAM [
111]. In low doses, the NO produced by this mechanism in tumor-bearing mice increased the angiogenesis of the tumor tissues, whereas the high doses led to tumor volume reduction and apoptotic tumor cell death [
111].
Immunotherapeutic approaches have demonstrated reprogramming M2 to M1 macrophages [
112,
113]. As stated earlier, induction of the innate immune response is initiated by activating the macrophage to M1-type, which produces NO/RNS, secrete TNF-α, IL-1β, and IL-6 with pro-inflammatory cytokines proteases such as MMP-9. Higher NO production activates downstream signaling pathways that perform a critical role in the cytotoxic activity of immune cells against tumor cells [
12,
114]. Furthermore, among other immune cells, NO synthesis in NK cells was shown to regulate their tumoricidal activity to some extent [
115].
2.1. iNOS Inhibitors
In colon, breast, gastric, hepatocellular carcinoma, melanoma, ovarian, leukemia, gastric, prostate, esophageal, and cervical cancers, high iNOS expression has correlated relatively well with poor patient survival [
116,
117]. Thus, iNOS expression may be used as a biomarker of poor patient prognosis and perhaps survival [
118,
119]. In contrast, a favorable prognosis has been associated with high iNOS expression in ovarian [
120] and non-small cell lung cancers [
121].
When cell lines expressing high levels of iNOS, such as that of triple-negative breast cancers, were treated with 1400W, which is a highly selective iNOS inhibitor, or L-NAME, which is a relatively selective eNOS inhibitor, or L-NMMA, which is pan-NOS inhibitor, they all reduced cell proliferation, migration, and mammosphere formation [
122]. In a xenograft model of TNBC, treatment of mice with L-NAME, and L-NMMA significantly reduced tumor growth [
122]. Administration of AG, another iNOS specific inhibitor to athymic nude mice bearing TNBC xenografts, abated tumor growth and metastatic burden [
123]. Further, the growth of glioma [
124] or melanoma [
125] cells in xenografts was significantly reduced when iNOS was silenced in these cells before they were implanted. The overarching data from all of these studies is the observation that the enhanced growth of the iNOS-overexpressing tumors appears to be due to enhanced angiogenesis, reviewed in [
3].
iNOS deficient mice exhibited enhanced M1 macrophage polarization with no significant effects on M2 macrophages. L-NIL, an iNOS selective inhibitor, significantly enhanced M1 macrophage polarization in cell cultures from wild-type (WT) mice. Whereas the NO donor SNAP, suppressed M1 macrophage differentiation in WT and
iNOS−/− cell cultures [
126].
5.2. NO and Curcumin: A Natural Dietary Compound
Natural dietary compounds modified or formulated as nanoparticles induce iNOS in TAMs and show promise in reprogramming M2 to M1 states to produce antitumor effects [
127]. A curcumin formulation containing two additional natural polyphenols, aptly named TriCurin, produced repolarization of M2 TAM that had higher Arg-1 expression into the M1 TAM population with higher iNOS expression. The underlying mechanism appears to be the suppression of activated STAT3 in M2 type TAM, which causes activation of STAT1, leading to the M1 phenotype. Co-activated transcription factors STAT1 and NF-κB initiate the expression of the iNOS and NO in M1 cells leading to tumor elimination [
128,
129]. Of note, these M1 phenotypes are low-IL10 and high-IL-12 and showed anti-glioblastoma activity [
130]. Specifically, for TriCurin, the M1 TAM-derived IL-12 that was induced was responsible for the recruitment of NK cells and cytotoxic T lymphocytes, leading to the reduction in cervical cancer cells in xenograft tumors [
129].
2.3. NO and Immunomodulation with microRNAs
MicroRNA-mediated regulation modulates macrophage states to the M2 or M1 phenotype [
131]. Delivery with nanomaterials has been developed wherein a marked increase in iNOS expression occurs for inducing or reprogramming the TAMs to M1-state or repolarization from M2- to M1-state, which provides an antitumor immune response (
Table 1). For example, Zhang et al. [
128] designed lipid-coated calcium phosphonate nanoparticles which were further conjugated with mannose for specific delivery of miR155 to TAMs, which altered phenotype successfully from pro-tumor M2-like TAMs to antitumor M1-like TAMs, and therefore produced a potent antitumor immune response and inhibiting tumor growth, reviewed in [
132]. Layered double hydroxides NPs that are miR-155 loaded are taken up by TAMs, increase their iNOS expression, decrease the expression level of phosphorylated STAT3 and ERK1/2 and activate NF-κB expression. Combined therapies showed improvement also, for example, these nanoparticles with carboplatin improved TC-1 tumor recession in animal models and prolonged overall survival [
133]. Similarly, Parayath et al. [
134] developed a CD44 targeting hyaluronic acid-poly(ethylenimine) (HA-PEI)-based nanoparticle for the delivery of miR-125b to peritoneal macrophages to promote M1-like TAMs activation in the lungs. In vivo results found a more than 6-fold increase in the ratio of M1 to M2 TAMs and a 300-fold increase in iNOS to Arg-1 in TAMs after treatment with HA-PEI-125b nanoparticles. The continued success of inducing M1-like TAMs polarization opens many avenues in anticancer immunotherapy via NO and enhancement of ROS release (
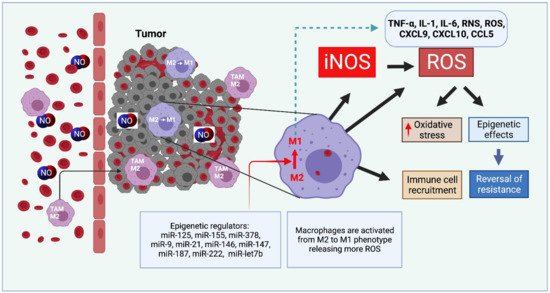
Figure 1).
Figure 1. Exogenous NO or iNOS-derived NO modulate the macrophage status. M2-type macrophages may be re-polarized into M1 phenotype via regulatory miRNA. ROS-generated oxidative stress may produce a cytocidal profile and reverse tumor progression.
2.4. NO-Releasing Nanoparticles
Synthetic compounds and various carriers also hold promise considering that years of iNOS focus have created avenues to mimic iNOS/ NO activity in various ways [
1]. Nanoparticles are demonstrated to preferentially accumulate in macrophages after systemic administration [
135]. Nanoparticles made to release NO produce cytotoxicity depending on the nanocarrier’s chemical nature, the concentration of NO released and the cell type. Low concentrations of NO released will have a proliferative effect on tumor cells, whereas high NO flux is expected to have toxic effects. There is considerable interest in identifying delivery methods to modulate TAM polarization for cancer treatment. TAMs overexpress the macrophage mannose receptor and therefore, mannose functionalized nanoparticles are used for recognition and internalization [
136]. Affinity to TAMs was improved via a mannose- conjugate modified on lipid-coated calcium phosphonate nanoparticles which delivered miRNA into TAMs in vitro [
137]. Some NO-releasing nanoparticles and materials with potential use in cancer treatment are presented in
Table 1 and principal actions are discussed. Reviews of various types of nanoparticles and effects on TME are found in, [
135,
138,
139].
Certain NO-releasing nanoparticles have been designed for photo-release. Nanoparticles of supramolecular assemblies of cyclodextrin-based polymer contain a NO photo donor and a fluorophore/photochrome dyad with an average size of 30 nm releases NO by light input [
140]. Photogenerated NO in human melanoma cancer cells showed cytotoxicity with light stimulation and low levels of cytotoxicity in the dark. Similarly, internalized nanoparticles NONOate into the endosomes and lysosomes of a cell, and cytotoxicity from NO-mediated apoptosis was found with NONOate modified silica nanoparticles which produced high NO flux [
141]. However, the desired sustained NO release akin to endogenous production by iNOS/ NO was not obtained.
Gold nanoparticles with 2-mercapto-5-nitrobenzimidazole also photo release NO and a low dose has similar antitumor and cytotoxicity effects as cisplatin in HeLa, Siha (cervical cancer cell lines), MCF-7 (breast cancer cell lines), and A549 (lung cancer lines); wherein 80% lower dose of Gold nanoparticles was found to produce cytotoxicity as that of 10 g/mL of cisplatin [
142].
Table 1. Nanoparticles releasing NO or inducing iNOS and effect on macrophages or TAM.
| Nanoparticles and Effect on iNOS or NO |
Model System or Cell Type |
Effect |
Reference |
| CD44 coated HA-PEI based NPs, miR-125b loaded, iNOS increased |
Naïve and KRAS/p53 double mutant nonsmall cell lung cancer (NSCLC) mouse model |
Specifically target peritoneal macrophages which reprogram lung TAMs into M1 type |
[143] |
Layered double hydroxides NPs, miR-155 loaded, acidity sensitive, taken up by TAM
iNOS increased |
TC-1 mouse tumor model
Uptake by TAM
Repolarize TAM into M1 |
Synergistic enhancement of therapeutic effects with programmed cell death-1 antibody (α-PD-1) antibody |
[133] |
Lipid-coated calcium
phosphonate, miR-155
conjugated mannose,
iNOS increased |
S180 mouse sarcoma model |
Repolarize M2 into M1 TAMs
Significant antitumor effect |
[137] |
Gold nanoparticles,
Photo release of NO |
HeLa |
Low doses of Gold nanoparticles were found to produce cytotoxicity as that of 10 g/mL of cisplatin |
[142] |
Poly(D,L-lactic-co-glycolic) acid (PLGA), loaded with ruthenium nitrosyl compounds,
NO releasing upon light irradiation |
Melanoma B16-F10 cells |
In vitro cytotoxicity assays showed cell death |
[144] |
Cyclodextrin and NO
photorelease by a donor |
HeLa, Melanoma,
A431- Human squamous carcinoma,
Melanoma |
Phototoxicity
cell mortality |
[145]
[146]
[147] |
Polymeric,
NO-releasing |
BE(2)-C, Neuroblastoma cell line |
Cisplatin in combination with nanoparticles produced synergistic cytotoxicity |
[148] |
| 4-arm branched polymer, NO-releasing |
Human head and neck cancer cell line human breast cancer cell lines |
Improved cell mortality |
[149] |
| Liposome, NO-releasing |
Breast cancer cell lines MDA-MB-231 and MDAMB-468 |
Improved cell mortality |
[150] |
In addition to NONOates, S-nitrosothiols (RSNOs) such as S-nitrosoglutathione (GSNO) were encapsulated into polymeric nanoparticles. GSNO was incorporated into polymeric nanoparticles consisting of diblock copolymers, which extended the RSNO stability. Because the combination of NO donors with classical chemotherapy agents is of considerable interest, GSNO-containing polymeric nanoparticles and cisplatin were used in in vitro experiments [
148]. NO-polymeric nanoparticles showed enhanced NO stability in aqueous media, were non-toxic and could efficiently release NO intracellularly [
148]. Neuroblastoma cell lines treated with GSNO-containing polymeric nanoparticles followed by cisplatin provided sensitization of cells and lower IC50 of cisplatin. NONOate-multiarm polymer nanocarriers to tumor-bearing nude mice inhibited tumor growth and extended the average survival of the animals in 7 weeks compared with intravenous administration of the classical NO-donor prodrug JS-K, and owed to a steady NO release profile. In vivo models may be reexamined for effects on macrophages [
148].
It is important to note that nanoparticle-based studies on the release of NO and the effects on the TME or TAMs should use appropriate reference treatment conditions for accurate comparisons of the efficiency of nanoparticle-induced effects. Nanoparticle-induced macrophage programming effects are generally compared to small molecule-induced effects. Some anticancer drugs may suppress immune activity within tumors and promote tumor growth and continue to be used in the clinical landscape. Therefore, it is imperative to compare the effect of nanoparticle-treated macrophages to biomolecule-treated macrophages, for example, with biomolecules such as IFN-γ, IL-4, IL-10, or LPS, or drug-treated macrophages for advancing the field immunotherapy based on NO.