Cold stress, including freezing stress and chilling stress, is one of the major environmental factors that limit the growth and productivity of plants. As a temperate dicot model plant species, Arabidopsis develops a capability to freezing tolerance through cold acclimation. The past decades have witnessed a deep understanding of mechanisms underlying cold stress signal perception, transduction, and freezing tolerance in Arabidopsis. In contrast, a monocot cereal model plant species derived from tropical and subtropical origins, rice, is very sensitive to chilling stress and has evolved a different mechanism for chilling stress signaling and response.
- cold stress
- signal perception
- Arabidopsis
- rice
1. Cold Stress Perceptions at Plasma Membrane (PM) in Arabidopsis and Rice
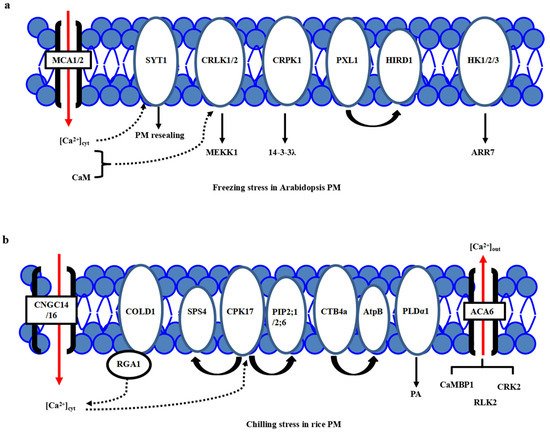
2. Cold Stress Signal Transduction Mechanisms in Arabidopsis and Rice
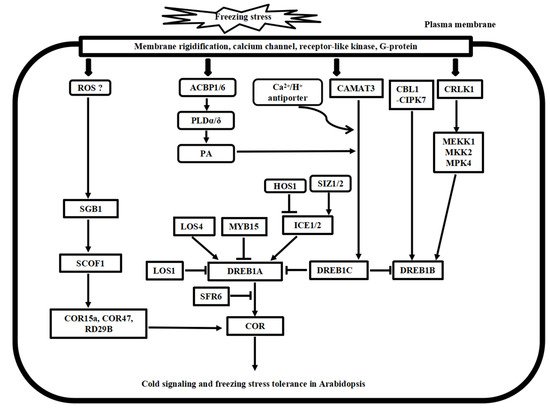
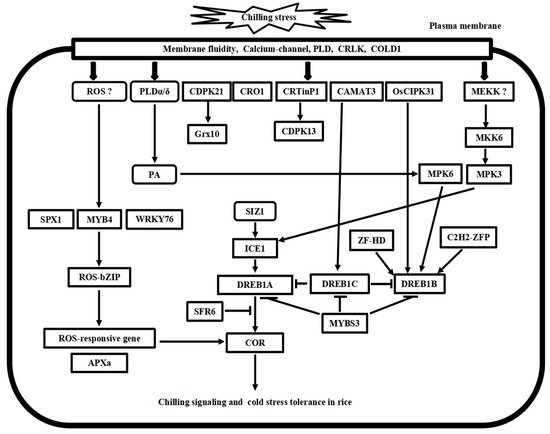
2.1. Calcium Signaling
2.2. Phospholipid Signaling
2.3. MAPK Cascade Signaling
2.4. ROS Signaling
This entry is adapted from the peer-reviewed paper 10.3390/plants10091864
References
- Murata, N.; Los, D.A. Membrane fluidity and temperature perception. Plant. Physiol. 1997, 115, 875–879.
- Sharma, A.; Isogai, M.; Yamamoto, T.; Sakaguchi, K.; Hashimoto, J.; Komatsu, S. A novel interaction between calreticulin and ubiquitin-like nuclear protein in rice. Plant. Cell Physiol. 2004, 45, 684–692.
- Penfield, S. Temperature perception and signal transduction in plants. New Phytol. 2008, 179, 615–628.
- McClung, C.R.; Davis, S.J. Ambient thermometers in plants: From physiological outputs towards mechanisms of thermal sensing. Curr. Biol. 2010, 20, R1086–R1092.
- Knight, M.R.; Knight, H. Low-temperature perception leading to gene expression and cold tolerance in higher plants. New Phytol. 2012, 195, 737–751.
- Monroy, A.F.; Dhindsa, R.S. Low-temperature signal transduction: Induction of cold acclimation-specific genes of alfalfa by calcium at 25 degrees c. Plant. Cell 1995, 7, 321–331.
- Knight, H.; Trewavas, A.J.; Knight, M.R. Cold calcium signaling in arabidopsis involves two cellular pools and a change in calcium signature after acclimation. Plant. Cell 1996, 8, 489–503.
- Kiegle, E.; Moore, C.A.; Haseloff, J.; Tester, M.A.; Knight, M.R. Cell-type-specific calcium responses to drought, salt and cold in the arabidopsis root. Plant. J. 2000, 23, 267–278.
- Mori, K.; Renhu, N.; Naito, M.; Nakamura, A.; Shiba, H.; Yamamoto, T.; Suzaki, T.; Iida, H.; Miura, K. Ca(2+)-permeable mechanosensitive channels mca1 and mca2 mediate cold-induced cytosolic ca(2+) increase and cold tolerance in arabidopsis. Sci. Rep. 2018, 8, 550.
- Ma, Y.; Dai, X.; Xu, Y.; Luo, W.; Zheng, X.; Zeng, D.; Pan, Y.; Lin, X.; Liu, H.; Zhang, D. Cold1 confers chilling tolerance in rice. Cell 2015, 160, 1209–1221.
- Cui, Y.; Lu, S.; Li, Z.; Cheng, J.; Hu, P.; Zhu, T.; Wang, X.; Jin, M.; Wang, X.; Li, L.; et al. Cyclic nucleotide-gated ion channels 14 and 16 promote tolerance to heat and chilling in rice. Plant. Physiol. 2020, 183, 1794–1808.
- Wang, J.; Ren, Y.; Liu, X.; Luo, S.; Zhang, X.; Liu, X.; Lin, Q.; Zhu, S.; Wan, H.; Yang, Y.; et al. Transcriptional activation and phosphorylation of oscngc9 confer enhanced chilling tolerance in rice. Mol. Plant. 2021, 14, 315–329.
- Yamazaki, T.; Kawamura, Y.; Minami, A.; Uemura, M. Calcium-dependent freezing tolerance in arabidopsis involves membrane resealing via synaptotagmin syt1. Plant. Cell 2008, 20, 3389–3404.
- Huang, R.; Zhao, J.; Liu, J.; Wang, Y.; Han, S.; Zhao, H. Genome-wide analysis and expression profiles of ntmc2 family genes in oryza sativa. Gene 2017, 637, 130–137.
- Jeon, J.; Kim, J. Arabidopsis response regulator1 and arabidopsis histidine phosphotransfer protein2 (ahp2), ahp3, and ahp5 function in cold signaling. Plant. Physiol. 2013, 161, 408–424.
- Jeon, J.; Kim, N.Y.; Kim, S.; Kang, N.Y.; Novak, O.; Ku, S.J.; Cho, C.; Lee, D.J.; Lee, E.J.; Strnad, M.; et al. A subset of cytokinin two-component signaling system plays a role in cold temperature stress response in arabidopsis. J. Biol. Chem. 2010, 285, 23371–23386.
- Yang, T.; Shad Ali, G.; Yang, L.; Du, L.; Reddy, A.S.; Poovaiah, B.W. Calcium/calmodulin-regulated receptor-like kinase crlk1 interacts with mekk1 in plants. Plant. Signal. Behav. 2010, 5, 991–994.
- Liu, Z.; Jia, Y.; Ding, Y.; Shi, Y.; Li, Z.; Guo, Y.; Gong, Z.; Yang, S. Plasma membrane crpk1-mediated phosphorylation of 14-3-3 proteins induces their nuclear import to fine-tune cbf signaling during cold response. Mol. Cell 2017, 66, 117–128.e5.
- Jung, C.G.; Hwang, S.G.; Park, Y.C.; Park, H.M.; Kim, D.S.; Park, D.H.; Jang, C.S. Molecular characterization of the cold- and heat-induced arabidopsis pxl1 gene and its potential role in transduction pathways under temperature fluctuations. J. Plant. Physiol. 2015, 176, 138–146.
- Kamrul Huda, K.M.; Akhter Banu, M.S.; Yadav, S.; Sahoo, R.K.; Tuteja, R.; Tuteja, N. Salinity and drought tolerant osaca6 enhances cold tolerance in transgenic tobacco by interacting with stress-inducible proteins. Plant. Physiol. Biochem. 2014, 82, 229–238.
- Almadanim, M.C.; Alexandre, B.M.; Rosa, M.T.; Sapeta, H.; Leitão, A.E.; Ramalho, J.C.; Lam, T.T.; Negrão, A.; Abreu, I.A.; Oliveira, M.M. Rice calcium-dependent protein kinase oscpk17 targets plasma membrane intrinsic protein and sucrose-phosphate synthase and is required for a proper cold stress response. Plant. Cell Environ. 2017, 40, 1197–1213.
- Zhang, Z.; Li, J.; Pan, Y.; Zhou, L.; Shi, H.; Zeng, Y.; Guo, H.; Yang, S.; Zheng, W.; Yu, J.; et al. Natural variation in ctb4a enhances rice adaptation to cold habitats. Nat. Commun. 2017, 8, 14788.
- Huo, C.; Zhang, B.; Wang, H.; Wang, F.; Liu, M.; Gao, Y.; Zhang, W.; Deng, Z.; Sun, D.; Tang, W. Comparative study of early cold-regulated proteins by two-dimensional difference gel electrophoresis reveals a key role for phospholipase dalpha1 in mediating cold acclimation signaling pathway in rice. Mol. Cell Proteom. 2016, 15, 1397–1411.
- Chinnusamy, V.; Zhu, J.K.; Sunkar, R. Gene regulation during cold stress acclimation in plants. Methods Mol. Biol. 2010, 639, 39–55.
- Chu, M.; Li, J.; Zhang, J.; Shen, S.; Li, C.; Gao, Y.; Zhang, S. Atcam4 interacts with a sec14-like protein, patl1, to regulate freezing tolerance in arabidopsis in a cbf-independent manner. J. Exp. Bot. 2018, 69, 5241–5253.
- Huang, C.; Ding, S.; Zhang, H.; Du, H.; An, L. Cipk7 is involved in cold response by interacting with cbl1 in arabidopsis thaliana. Plant. Sci. 2011, 181, 57–64.
- Doherty, C.J.; Van Buskirk, H.A.; Myers, S.J.; Thomashow, M.F. Roles for arabidopsis camta transcription factors in cold-regulated gene expression and freezing tolerance. Plant. Cell 2009, 21, 972–984.
- Catala, R.; Santos, E.; Alonso, J.M.; Ecker, J.R.; Martinez-Zapater, J.M.; Salinas, J. Mutations in the ca2+/h+ transporter cax1 increase cbf/dreb1 expression and the cold-acclimation response in arabidopsis. Plant. Cell 2003, 15, 2940–2951.
- Liu, Q.; Ding, Y.; Shi, Y.; Ma, L.; Wang, Y.; Song, C.; Wilkins, K.A.; Davies, J.M.; Knight, H.; Knight, M.R.; et al. The calcium transporter annexin1 mediates cold-induced calcium signaling and freezing tolerance in plants. EMBO J. 2021, 40, e104559.
- Yang, J.; Liu, S.; Ji, L.; Tang, X.; Zhu, Y.; Xie, G. Identification of novel oscml16 target proteins and differential expression analysis under abiotic stresses in rice. J. Plant. Physiol. 2020, 249, 153165.
- Saijo, Y.; Hata, S.; Kyozuka, J.; Shimamoto, K.; Izui, K. Over-expression of a single ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants. Plant. J. 2000, 23, 319–327.
- Saijo, Y.; Kinoshita, N.; Ishiyama, K.; Hata, S.; Kyozuka, J.; Hayakawa, T.; Nakamura, T.; Shimamoto, K.; Yamaya, T.; Izui, K. A ca(2+)-dependent protein kinase that endows rice plants with cold- and salt-stress tolerance functions in vascular bundles. Plant. Cell Physiol. 2001, 42, 1228–1233.
- Abbasi, F.; Onodera, H.; Toki, S.; Tanaka, H.; Komatsu, S. Oscdpk13, a calcium-dependent protein kinase gene from rice, is induced by cold and gibberellin in rice leaf sheath. Plant. Mol. Biol. 2004, 55, 541–552.
- Komatsu, S.; Yang, G.; Khan, M.; Onodera, H.; Toki, S.; Yamaguchi, M. Over-expression of calcium-dependent protein kinase 13 and calreticulin interacting protein 1 confers cold tolerance on rice plants. Mol. Genet. Genom. 2007, 277, 713–723.
- Liu, Y.; Xu, C.; Zhu, Y.; Zhang, L.; Chen, T.; Zhou, F.; Chen, H.; Lin, Y. The calcium-dependent kinase oscpk24 functions in cold stress responses in rice. J. Integr. Plant. Biol. 2018, 60, 173–188.
- Xiang, Y.; Huang, Y.; Xiong, L. Characterization of stress-responsive cipk genes in rice for stress tolerance improvement. Plant. Physiol. 2007, 144, 1416–1428.
- Kim, K.N.; Lee, J.S.; Han, H.; Choi, S.A.; Go, S.J.; Yoon, I.S. Isolation and characterization of a novel rice ca2+-regulated protein kinase gene involved in responses to diverse signals including cold, light, cytokinins, sugars and salts. Plant. Mol. Biol. 2003, 52, 1191–1202.
- Vaultier, M.N.; Cantrel, C.; Vergnolle, C.; Justin, A.M.; Demandre, C.; Benhassaine-Kesri, G.; Cicek, D.; Zachowski, A.; Ruelland, E. Desaturase mutants reveal that membrane rigidification acts as a cold perception mechanism upstream of the diacylglycerol kinase pathway in arabidopsis cells. FEBS Lett. 2006, 580, 4218–4223.
- Li, W.; Li, M.; Zhang, W.; Welti, R.; Wang, X. The plasma membrane-bound phospholipase ddelta enhances freezing tolerance in arabidopsis thaliana. Nat. Biotechnol. 2004, 22, 427–433.
- Rajashekar, C.B.; Zhou, H.E.; Zhang, Y.; Li, W.; Wang, X. Suppression of phospholipase dalpha1 induces freezing tolerance in arabidopsis: Response of cold-responsive genes and osmolyte accumulation. J. Plant. Physiol. 2006, 163, 916–926.
- Tan, W.J.; Yang, Y.C.; Zhou, Y.; Huang, L.P.; Xu, L.; Chen, Q.F.; Yu, L.J.; Xiao, S. Diacylglycerol acyltransferase and diacylglycerol kinase modulate triacylglycerol and phosphatidic acid production in the plant response to freezing stress. Plant. Physiol. 2018, 177, 1303–1318.
- Chen, Q.F.; Xiao, S.; Chye, M.L. Overexpression of the arabidopsis 10-kilodalton acyl-coenzyme a-binding protein acbp6 enhances freezing tolerance. Plant. Physiol. 2008, 148, 304–315.
- Du, Z.Y.; Chen, M.X.; Chen, Q.F.; Gu, J.D.; Chye, M.L. Expression of arabidopsis acyl-coa-binding proteins atacbp1 and atacbp4 confers pb(ii) accumulation in brassica juncea roots. Plant. Cell Env. 2015, 38, 101–117.
- Li, Q.; Zheng, Q.; Shen, W.; Cram, D.; Fowler, D.B.; Wei, Y.; Zou, J. Understanding the biochemical basis of temperature-induced lipid pathway adjustments in plants. Plant. Cell 2015, 27, 86–103.
- Ariizumi, T.; Kishitani, S.; Inatsugi, R.; Nishida, I.; Murata, N.; Toriyama, K. An increase in unsaturation of fatty acids in phosphatidylglycerol from leaves improves the rates of photosynthesis and growth at low temperatures in transgenic rice seedlings. Plant. Cell Physiol. 2002, 43, 751–758.
- Tovuu, A.; Zulfugarov, I.S.; Wu, G.X.; Kang, I.S.; Kim, C.; Moon, B.Y.; An, G.; Lee, C.H. Rice mutants deficient in omega-3 fatty acid desaturase (fad8) fail to acclimate to cold temperatures. Plant. Physiol. Biochem. 2016, 109, 525–535.
- Zheng, G.; Li, L.; Li, W. Glycerolipidome responses to freezing- and chilling-induced injuries: Examples in arabidopsis and rice. BMC Plant. Biol. 2016, 16, 70.
- Hamel, L.P.; Nicole, M.C.; Sritubtim, S.; Morency, M.J.; Ellis, M.; Ehlting, J.; Beaudoin, N.; Barbazuk, B.; Klessig, D.; Lee, J.; et al. Ancient signals: Comparative genomics of plant mapk and mapkk gene families. Trends Plant. Sci. 2006, 11, 192–198.
- Kovtun, Y.; Chiu, W.L.; Tena, G.; Sheen, J. Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants. Proc. Natl. Acad. Sci. USA 2000, 97, 2940–2945.
- Teige, M.; Scheikl, E.; Eulgem, T.; Doczi, R.; Ichimura, K.; Shinozaki, K.; Dangl, J.L.; Hirt, H. The mkk2 pathway mediates cold and salt stress signaling in arabidopsis. Mol. Cell 2004, 15, 141–152.
- Kim, S.H.; Kim, H.S.; Bahk, S.; An, J.; Yoo, Y.; Kim, J.Y.; Chung, W.S. Phosphorylation of the transcriptional repressor myb15 by mitogen-activated protein kinase 6 is required for freezing tolerance in arabidopsis. Nucleic Acids Res. 2017, 45, 6613–6627.
- Zhao, C.; Wang, P.; Si, T.; Hsu, C.C.; Wang, L.; Zayed, O.; Yu, Z.; Zhu, Y.; Dong, J.; Tao, W.A.; et al. Map kinase cascades regulate the cold response by modulating ice1 protein stability. Dev. Cell 2017, 43, 618–629.e5.
- Li, H.; Ding, Y.; Shi, Y.; Zhang, X.; Zhang, S.; Gong, Z.; Yang, S. Mpk3- and mpk6-mediated ice1 phosphorylation negatively regulates ice1 stability and freezing tolerance in arabidopsis. Dev. Cell 2017, 43, 630–642.e4.
- Xie, G.; Kato, H.; Imai, R. Biochemical identification of the osmkk6-osmpk3 signalling pathway for chilling stress tolerance in rice. Biochem. J. 2012, 443, 95–102.
- Zhang, Z.; Li, J.; Li, F.; Liu, H.; Yang, W.; Chong, K.; Xu, Y. Osmapk3 phosphorylates osbhlh002/osice1 and inhibits its ubiquitination to activate ostpp1 and enhances rice chilling tolerance. Dev. Cell 2017, 43, 731–743.e5.
- Vogel, J.T.; Zarka, D.G.; Van Buskirk, H.A.; Fowler, S.G.; Thomashow, M.F. Roles of the cbf2 and zat12 transcription factors in configuring the low temperature transcriptome of arabidopsis. Plant. J. 2005, 41, 195–211.
- Shi, H.; Ye, T.; Zhong, B.; Liu, X.; Jin, R.; Chan, Z. Athap5a modulates freezing stress resistance in arabidopsis through binding to ccaat motif of atxth21. New Phytol. 2014, 203, 554–567.
- van Buer, J.; Cvetkovic, J.; Baier, M. Cold regulation of plastid ascorbate peroxidases serves as a priming hub controlling ros signaling in arabidopsis thaliana. BMC Plant. Biol. 2016, 16, 163.
- Park, J.H.; Lee, E.S.; Chae, H.B.; Paeng, S.K.; Wi, S.D.; Bae, S.B.; Thi Phan, K.A.; Lee, S.Y. Disulfide reductase activity of thioredoxin-h2 imparts cold tolerance in arabidopsis. Biochem. Biophys. Res. Commun. 2021, 568, 124–130.
- Yun, K.Y.; Park, M.R.; Mohanty, B.; Herath, V.; Xu, F.; Mauleon, R.; Wijaya, E.; Bajic, V.B.; Bruskiewich, R.; de Los Reyes, B.G. Transcriptional regulatory network triggered by oxidative signals configures the early response mechanisms of japonica rice to chilling stress. BMC Plant. Biol. 2010, 10, 16.
- Cheng, C.; Yun, K.Y.; Ressom, H.W.; Mohanty, B.; Bajic, V.B.; Jia, Y.; Yun, S.J.; de los Reyes, B.G. An early response regulatory cluster induced by low temperature and hydrogen peroxide in seedlings of chilling-tolerant japonica rice. BMC Genom. 2007, 8, 175.
- Zhang, J.; Luo, W.; Zhao, Y.; Xu, Y.; Song, S.; Chong, K. Comparative metabolomic analysis reveals a reactive oxygen species-dominated dynamic model underlying chilling environment adaptation and tolerance in rice. New Phytol. 2016, 211, 1295–1310.
- Huang, J.; Sun, S.-J.; Xu, D.-Q.; Yang, X.; Bao, Y.-M.; Wang, Z.-F.; Tang, H.-J.; Zhang, H. Increased tolerance of rice to cold, drought and oxidative stresses mediated by the overexpression of a gene that encodes the zinc finger protein zfp245. Biochem. Biophys. Res. Commun. 2009, 389, 556–561.
- Sato, Y.; Masuta, Y.; Saito, K.; Murayama, S.; Ozawa, K. Enhanced chilling tolerance at the booting stage in rice by transgenic overexpression of the ascorbate peroxidase gene, osapxa. Plant. Cell Rep. 2011, 30, 399–406.
- Wang, X.; Zou, B.; Shao, Q.; Cui, Y.; Lu, S.; Zhang, Y.; Huang, Q.; Huang, J.; Hua, J. Natural variation reveals that ossap16 controls low-temperature germination in rice. J. Exp. Bot. 2018, 69, 413–421.