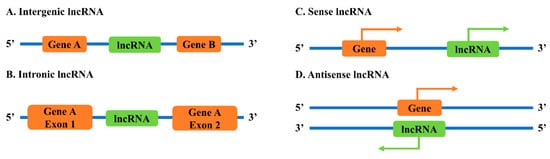
2.2. Neighborhood Antisense lncRNAs to Sense Genes
An antisense transcript refers to a transcript generated from the opposite strand to a protein-coding or sense strand [
17]. The existence of antisense transcription is very common across different transcriptomes and these antisense transcripts are usually co-transcribed with their sense transcripts [
18]. It has been demonstrated that more than 60% of the sense transcripts have antisense partners [
17,
19,
20]. Antisense transcripts have been reported to have multiple functions in the regulation of gene expression, which have further impacts on human health and diseases [
21,
22].
lncRNAs have been identified to be one common type of antisense transcripts of coding genes and are involved in the regulation of function and expression of their neighborhood coding genes [
23,
24]. These antisense lncRNAs also have several special characteristics, which are similar to mRNAs. There are several structural similarities between the lncRNAs and mRNAs, where both can be multi-exonic, 5′-capped, and 3′-polyadenylated [
25]. The processing of lncRNAs is also similar to mRNAs, where the RNA polymerase II is responsible for the synthesis and both RNAs need to be spliced [
21]. The transcriptional activity of lncRNAs can also be regulated by special DNA elements in promoters and enhancers, where promoters can have different locations [
26]. In some special cases, both the sense and antisense genes can be controlled by a same bidirectional promoter, which causes the co-expression of the sense and antisense gene pair [
27].
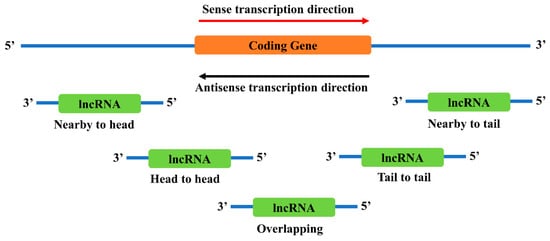
There are several locations near a coding gene where antisense lncRNAs are likely to exist (Figure 2). However, it is still not clear how different locations affect the function of lncRNAs. Recent studies have found that these neighborhood lncRNAs might affect their nearby coding genes in multiple ways, including direct regulation of the coding genes or indirect involvement in the functional regulation of the coding genes.
Figure 2. Genomic locations of antisense neighborhood lncRNAs to a sense coding gene. Depending on the relative locations towards the coding gene, neighborhood lncRNAs are categorized into several types: nearby to head, the 5’ end of the lncRNA is close to the 5’ end of the coding gene; head to head, the 5’ end of both genes are aligned together; overlapping, the antisense lncRNA overlaps with the sense coding gene; tail to tail, the 3’ end of both genes are aligned together; and nearby to tail, the 3’ end of the lncRNA is close to the 3’ end of the coding gene.
2.3. Neighborhood Antisense lncRNAs to TFs
Along with the advancements in the discovery of lncRNAs, there is an increasing amount of studies trying to identify lncRNAs that have special relationships with coding genes. These relationships include but are not limited to correlations in expression, spatial localization, and function [
28,
29,
30]. There is increasing evidence showing that lncRNAs, including TF neighborhood antisense lncRNAs, are involved in the expression or functions of TFs in gene regulation [
31]. In this section, several TF-lncRNA pairs are summarized and discussed (
Table 1).
Table 1. Summary of TF-neighborhood lncRNA pairs.
GATA3-AS1 is a divergent lncRNA gene sharing a promoter region with the TF gene
GATA3, encoding a master regulator in T helper 2 cell functions. In one study performed by Gibbons et al., GATA3-AS1 was found to control the expression of
GATA3 gene as well as two interleukin genes,
IL5 and
IL13. Specifically, depletion of GATA3-AS1 resulted in a reduction in GATA3, IL5, and IL13 mRNA and protein levels, along with decreased T helper 2 cell polarization. This regulatory function of GATA3-AS1 was accomplished through the interaction with histone-modifying enzymes and the alteration of histone codes to its own gene locus, which is the shared promoter region of
GATA3 [
32].
GA-binding protein subunit beta-1 (GABPB1) is a TF known to regulate the transcription of various genes related to antioxidation process, including Peroxiredoxin 5 (
PRDX5) [
35]. lncRNA
GABPB1-AS1 gene is an antisense RNA gene of the
GABPB1 gene. GABPB1 protein was previously reported to respond to chemical-induced cellular stress [
36]. A recent study showed that GABPB1-AS1 was able to affect the functions of GABPB1 by modulating the translation process of GABPB1 in HepG2 cells, an
in vitro model for hepatocellular carcinoma (HCC). The upregulation of GABPB1-AS1 inhibited the translation of GABPB1, which further reduced the expression of peroxidase genes, including PRDX5, and accumulation of reactive oxygen species and lipid peroxidation in the cells [
33].
Yin Yang 1 (YY1) is an ubiquitously expressed TF involved in the regulation of cell proliferation and differentiation [
37]. In muscle cells or myoblasts, YY1 regulates genes involved in cell proliferation and differentiation by interacting with histone methyltransferase complex and altering histone markers on target genes [
38].
Linc-YY1 gene is a divergently transcribed lncRNA gene located upstream of the
YY1 coding gene. The employment of linc-YY1 loss- and gain-of-function assays in a mouse myoblast cell line showed that linc-YY1 was important in the myogenic differentiation and muscle regeneration processes, and modulation of
linc-YY1 also affected YY1 target genes. This function of linc-YY1 was accomplished through interacting with YY1 directly and eviction of YY1/Polycomb repressive complex, a complex mediating histone methylation from promoter regions of target genes [
34].
All these studies supported the idea that lncRNAs, especially neighborhood lncRNAs of TFs, might be involved in the function of these TFs.
2.4. Genomic Locations and Structures of HNF1α-AS1 and HNF4α-AS1
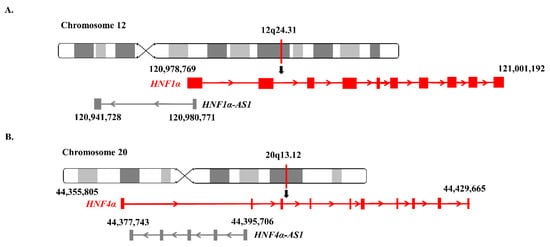
HNF1α-AS1, a neighbor antisense lncRNA gene of the human HNF1α gene, is located at human Chromosome 12 with a length of 39.04 kb, containing two exons and one intron. The coding gene HNF1α and noncoding gene HNF1α-AS1 formed a typical TF-lncRNA pair (Figure 3A).
Figure 3. Genomic locations of HNF1α, HNF1α-AS1, HNF4α, and HNF4α-AS1: (A) locations of HNF1α and HNF1α-AS1 on human chromosome 12; and (B) locations of HNF4α and HNF4α-AS1 on human chromosome 20.
HNF4α-AS1, a neighbor antisense lncRNA gene of the human HNF4α gene, is located at human Chromosome 20 with a length of 17.96 kb, containing four exons and three introns. The HNF4α and HNF4α-AS1 genes also form a typical pair of coding and neighborhood antisense noncoding genes (Figure 3B).
The coding potentials of HNF1α and HNF1α-AS1 as well as HNF4α and HNF4α-AS1 were analyzed by the Coding Potential Calculator (
http://cpc2.cbi.pku.edu.cn) [
39]. The analysis results (
Table 2) indicate that both HNF1α-AS1 and HNF4α-AS1 have low coding probabilities in comparison to HNF1α and HNF4α, which confirmed the noncoding features of HNF1α-AS1 and HNF4α-AS1.
Table 2. The coding potentials of HNF1α-AS1 and HNF4α-AS1.
The RNA transcripts of both coding genes of
HNF1α and
HNF4α and noncoding genes of
HNF1α-AS1 and
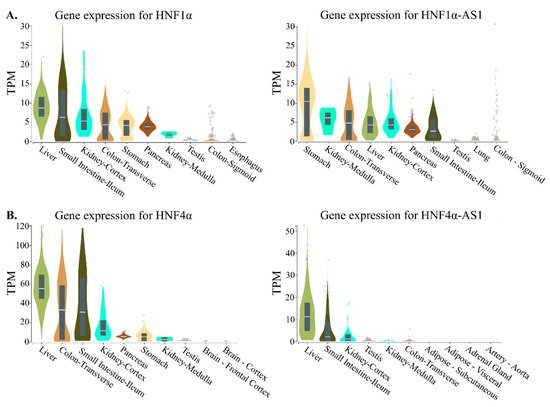
HNF4α-AS1 show tissue-specific distributions in normal human tissues. To compare the expression patterns between the coding and noncoding genes, the top 10 tissues expressing HNF1α, HNF1α-AS1, HNF4α, and HNF4α-AS1 in human are summarized and compared. The tissue distribution pattern of HNF1α mRNA was retrieved from the RNA-Seq Expression Data GTEx in 53 tissues from 570 donors and shows relatively higher expression levels in the stomach, small intestine, colon, liver, pancreas, and kidney, the major organs in the gastrointestinal (GI) tract (
Figure 4) [
40]. A similar pattern in the GI tract organs is also found for HNF1α-AS1 (
Figure 4A), indicating that HNF1α-AS1 is expressed in the organs where HNF1α is expressed. These results may suggest that HNF1α-AS1 is possibly involved in the regulatory function of HNF1α in the GI tract organs. Similar tissue distribution patterns are also found between HNF4α and HNF4α-AS1 (
Figure 4B). These similarities in tissue distribution might suggest functional connections between HNF1α and HNF1α-AS1 as well as between HNF4α and HNF4α-AS1.
Figure 4. Top 10 tissues expressing HNF1α, HNF1α-AS1, HNF4α, and HNF4α-AS1 in human: (
A) tissue expression patterns of HNF1α and HNF1α-AS1; and (
B) tissue expression patterns of HNF4α and HNF4α-AS1. Data are from GTEx database (
https://www.gtexportal.org/home/). TPM: transcripts per million.
In addition to the chromatin locations and tissue distribution, the structures of lncRNAs are also important for their function and mechanism of action. RNA molecules have been shown to adopt higher-order tertiary interactions [
41]. Even though the relationships between lncRNA structure and functions are still not fully understood yet, identification of structural domains, which mediate the interactions between lncRNAs and other molecules, is critical for the characterization of lncRNA functions. The secondary structures of HNF1α-AS1 is predicated using RNAfold program (
http://rna.tbi.univie.ac.at//cgi-bin/RNAWebSuite/RNAfold.cgi). The secondary structure of HNF4α-AS1 is not predicated as the sequence length of HNF4α-AS1 exceeds the upper limit of lncRNA FASTA input for the program.
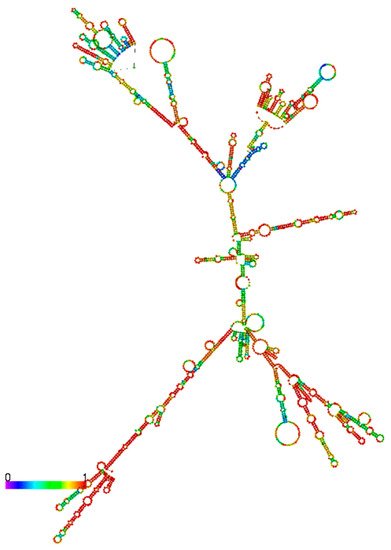
As shown in Figure 5, HNF1α-AS1 is able to form a stable secondary structure based on minimum free energy calculation. Several domains are also observed on the secondary structure of HNF1α-AS1. However, it is still not very clear what molecules interact with these domains and it is still largely unknown how they impact on the functions of HNF1α-AS1.
Figure 5. Predication of secondary structure of HNF1α-AS1 by RNAfold. The structure of HNF1α-AS1 is generated based on minimum free energy method. The colors indicate base-pair probabilities from low to high.
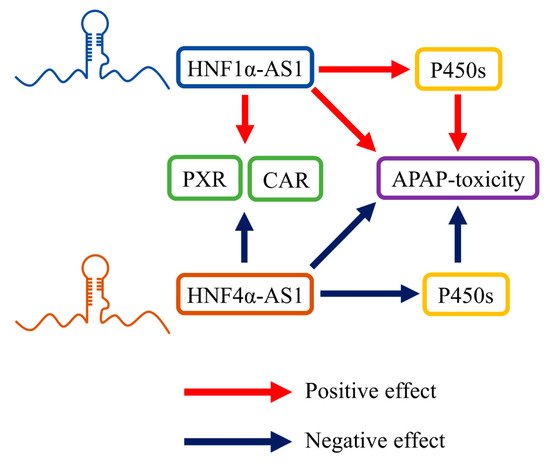
HNF1α and HNF4α are well-studied TFs involved in organ maturation, cell differentiation, and disease development. Both HNF1α and HNF4α can be detected during different stages of embryonic development and they participate in the development of multiple organs, including liver, colon, and pancreas [
42,
43,
44]. HNF1α and HNF4α are also regarded as master regulators of the metabolic functions in human. The target genes of HNF1α and HNF4α are involved in lipid metabolism, bile acid synthesis, lipoprotein metabolism, glucose metabolism, amino acid metabolism, and xenobiotic metabolism [
45,
46,
47,
48,
49]. Taking xenobiotic metabolism as an example, knockdown of HNF1α or HNF4α in mice or human primary hepatocytes led to downregulation of the mRNA expression of several cytochrome P450s (P450s), which play critical roles in metabolizing drugs [
49,
50]. In addition to normal physiological activities, HNF1α and HNF4α are also highly involved in several diseases. Genetic mutations in
HNF1α or
HNF4α gene are some of the most common causes of maturity-onset diabetes of the young (MODY), which is characterized by a non-insulin dependent form of diabetes in young people below the age of 25 [
51]. The dysfunction of pancreatic beta cells and impaired insulin secretion because of HNF1α or HNF4α mutations are believed to be the pathological mechanisms to this type of disease. In addition to MODY, HNF1α and HNF4α are involved in other type of diseases, including other metabolic diseases, inflammatory diseases, and cancer [
52,
53,
54,
55]. HNF1α and HNF4α are reported to play different roles based on cancer types. In HCC, the transduction of HNF1α and HNF4α was showed to suppress the growth of HepG2 and Huh7 cells
in vitro and to reduce the tumorigenicity of these cells after transplantation into mice, indicating a role of tumor suppressing of these TFs [
56]. However, in pancreatic cancer, the role of HNF1α is controversial based on the results from two reports. Luo and his colleagues reported that HNF1α was a possible tumor suppressing gene in pancreatic cancer [
57]. The results of immunohistochemistry showed that the level of HNF1α was significantly lower in pancreatic cancer tissues than normal pancreatic tissues. Furthermore, knockdown of HNF1α also led to increase proliferation and to decrease apoptosis in pancreatic cancer cell lines, which supported the conclusion that HNF1α plays a role in tumor suppressing. This conclusion was challenged by another report by Abel et al., in which the authors showed that overexpression of HNF1α increased the formation of pancreatic cancer stem cells and tumorsphere [
58]. In summary, HNF1α and HNF4α are two very important TFs in human physiologic functions and diseases.
Comparing to the well identified functions of HNF1α and HNF4α, the underlying molecular mechanisms of how HNF1α and HNF4α perform these functions is still not clearly understood yet. As TFs in nature, HNF1α and HNF4α can regulate their target genes through directly binding to their promoter regions. Other than direct binding, some other mechanisms of HNF1α and HNF4α in gene regulation are still elusive and need to be identified. lncRNAs are found to serve as cofactors of several important regulatory proteins and to be involved in their functions. However, whether lncRNAs are also involved in the function of HNF1α and HNF4α is still not fully understood.