Dysregulation of gap junction intercellular communication (GJIC) is recognized as one of the key hallmarks for identifying non-genotoxic carcinogens (NGTxC). Currently, there is a demand for in vitro assays addressing the gap junction hallmark, which would have the potential to eventually become an integral part of an integrated approach to the testing and assessment (IATA) of NGTxC. The scrape loading-dye transfer (SL-DT) technique is a simple assay for the functional evaluation of GJIC in various in vitro cultured mammalian cells and represents an interesting candidate assay. Out of the various techniques for evaluating GJIC, the SL-DT assay has been used frequently to assess the effects of various chemicals on GJIC in toxicological and tumor promotion research.
- carcinogenesis
- carcinogens
- gap junction intercellular communication
- scrape loading-dye transfer
1. Introduction
Chemical carcinogens can be classified into three main groups [1][2]: (1) ultimate carcinogens (chemicals with a direct action with the capacity to induce cancer without a previous metabolic activation), (2) procarcinogens (chemicals that need to be activated by metabolic activation to become ultimate carcinogens) and (3) co-carcinogens (chemical substances that cannot induce cancer when administered alone but can enhance the carcinogenic effect of other substances). From a toxicological and regulatory perspective, chemical carcinogens can be classified according to their prevailing mechanism as genotoxic carcinogens (GTxC), which include mutagenic or genotoxic agents inducing mutations and DNA damage by “errors of DNA repair” during initiation and eventually also progression stage. In contrast, NGTxCs (i.e., non-genotoxic carcinogens) represent agents whose carcinogenic activity does not depend on DNA damage but on various mechanisms altering cellular behavior during tumor promotion and progression stage [3]. In addition to an initiating agent being mutagenic, while a promoting agent is not mutagenic, there are other differences between the action of GTxC versus NGTxC. An initiating agent after repeated exposure in a small dosage or a single large exposure leads to carcinogenesis, in contrast to a promoting agent, which is not carcinogenic alone or when not exceeding a “threshold” limit. The duration and regularity of exposure rather than its intensity appear to be the most critical factors, as well as the absence of “anti-promotors”. An effect of an initiating carcinogen is irreversible and additive, whereas an effect of a promoting agent is reversible at the early stages [3].
The alternative, using in vitro testing methods and batteries, has already been established for GTxC, and some assays developed into OECD Test Guidelines [4]. Still, there are no available in vitro test guidelines addressing specifically human-relevant NGTxC [5]. To address the current lack of alternative testing tools and approaches, an OECD expert group developed an integrated approach to the testing and assessment (IATA) of chemical NGTxC [5][6]. Refined and structured in accordance with recognized cancer hallmarks and mechanistic knowledge, this IATA identified 13 key cancer hallmarks of NGTxC: (1) receptor binding and activation, including also hormone-mediated processes, and CYP P450 induction, (2) cell proliferation and (3) transformation, (4) GJIC (i.e., gap junction intercellular communication), (5) oxidative stress induction, (6) immunosuppression/immune evasion, (7) gene expression and cell signaling pathways, (8) increased resistance to apoptotic cell death, (9) pathogenic angiogenesis and neoangiogenesis, (10) genetic instability, (11) cellular senescence/telomerase, (12) invasion and metastasis and (13) epigenetic mechanisms [5][6]. These hallmarks are related to the key events occurring in the early to mid to later stages of the carcinogenic process. Based on this IATA framework and following the proposed assay evaluation criteria [5], appropriate tests, primarily in vitro assays, shall be identified and prioritized for further development and (pre)validation. The selected assay(s) will be targeted for validation needed for test guidelines and regulatory use. The representative standardized or commonly used tests (if available) addressing the key cancer hallmarks have recently been summarized, including the current status regarding their use in hazard assessment, availability of the test guidelines and their readiness level and eventually their inclusion into the OECD Test Guidelines Programme [5]. Cell-to-cell communication mediated through gap junction channels, i.e., GJIC, represents one of these essential key mechanisms for which there are currently no test guidelines or standardized tests [5].
GJIC is a fundamental biological cellular process in multi-cellular metazoan organisms that allows an exchange of various soluble ions and aqueous molecules between adjacent cells, allowing them to integrate multiple signals and coordinate their behavior in the tissues [7][8]. GJIC is a key mechanism for maintaining tissue homeostasis, and its dysregulation has been long recognized as a hallmark of NGTxC [9][5][6][10][8][11]. The inclusion of GJIC into the IATA of chemical NGTxC [5] has, thus, provided an incentive for evaluation, prioritization and further development of in vitro assays capable of addressing this specific hallmark, particularly with respect to the lack of existing test guidelines or candidate assays for GJIC hazard assessment within the OECD Test Guidelines Programme.
Among various techniques developed for in vitro assessment of GJIC, the SL-DT (i.e., scrape loading-dye transfer) assay has probably been most frequently used in multiple studies of toxicant or carcinogen effects on GJIC. This in vitro assay is applicable to various cell types and cell lines. However, most of the published data focusing on the chemical effects on GJIC were generated using a rat liver epithelial cell line WB-F344. Nevertheless, such information, which would be highly relevant for further prioritization of in vitro assays suitable to address the GJIC hallmark in the IATA for NGTxC, has yet to be systematically mapped and summarized.
2. GJIC as the Key Mechanism in Tissue Homeostasis
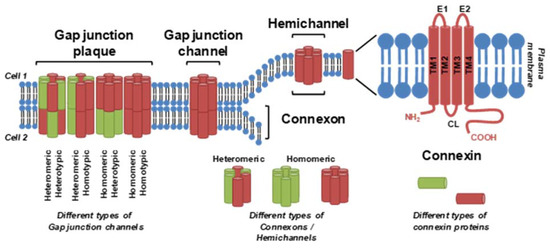
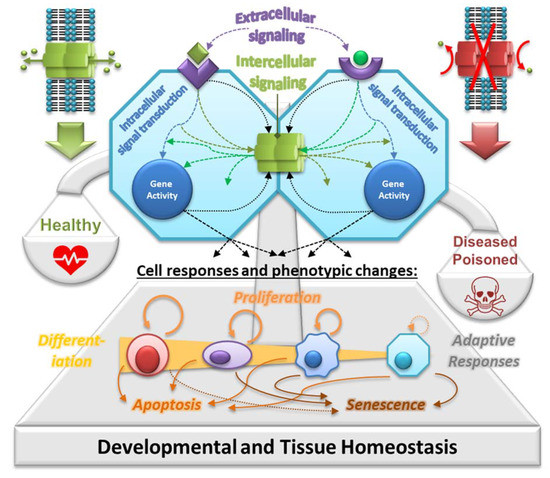
3. Cell Lines and Methods for In Vitro GJIC Assessment
The assays suitable for evaluating GJIC have been extensively reviewed, including discussions on their principles, applicability, advantages and disadvantages [13][17][18][19]. These assays can be principally divided into three major groups based on the technical approaches used for estimating GJIC capacity. Namely, there are assays based on the measurements of (a) electrical conductance (electrical coupling), such as the double whole-cell voltage-clamp (DWCV) technique, (b) endogenous metabolite transfer (metabolic cooperation assays, MCs) or (c) a fluorescent dye transfer (DT). The latter group involves a variety of techniques, such as fluorescence recovery after photobleaching (FRAP), local activation of fluorescent molecular probe (LAMP), microinjection (MI), scrape loading (SL) or preloading (Pre) and parachute (Par) assays.
4. Scrape Loading-Dye Transfer Using the WB-F344 Cell Line
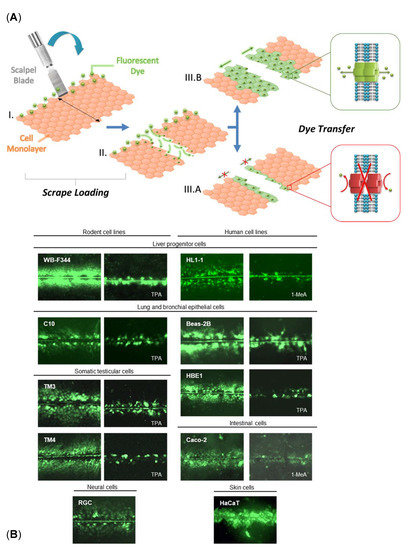
This entry is adapted from the peer-reviewed paper 10.3390/ijms22168977
References
- Baba, A.; Câtoi, C. Carcinogenesis. In Comparative Oncology; The Publishing House of the Romanian Academy: Bucharest, Romania, 2007.
- Bevan, R.J.; Harrison, P.T.C. Threshold and non-threshold chemical carcinogens: A survey of the present regulatory landscape. Regul. Toxicol. Pharmacol. 2017, 88, 291–302.
- Basu, A.K. DNA damage, mutagenesis and cancer. Int. J. Mol. Sci. 2018, 19, 970.
- OECD. Overview of the Set of OECD Genetic Toxicology Test Guidelines and Updates Performed in 2014–2015. 2017. Available online: https://www.oecd.org/officialdocuments/publicdisplaydocumentpdf/?cote=ENV-JM-MONO(2016)33/rev1&doclanguage=en (accessed on 16 June 2021).
- Jacobs, M.N.; Colacci, A.; Corvi, R.; Vaccari, M.; Aguila, M.C.; Corvaro, M.; Delrue, N.; Desaulniers, D.; Ertych, N.; Jacobs, A.; et al. Chemical carcinogen safety testing: OECD expert group international consensus on the development of an integrated approach for the testing and assessment of chemical non-genotoxic carcinogens. Arch. Toxicol. 2020, 1, 3.
- Jacobs, M.N.; Colacci, A.; Louekari, K.; Luijten, M.; Hakkert, B.C.; Paparella, M.; Vasseur, P. International regulatory needs for development of an IATA for non-genotoxic carcinogenic chemical substances. ALTEX 2016, 33, 359–392.
- Nielsen, M.S.; Nygaard Axelsen, L.; Sorgen, P.L.; Verma, V.; Delmar, M.; Holstein-Rathlou, N.-H. Gap junctions. Compr. Physiol. 2012, 2, 1981–2035.
- Zefferino, R.; Piccoli, C.; Di Gioia, S.; Capitanio, N.; Conese, M. Gap junction intercellular communication in the carcinogenesis hallmarks: Is this a phenomenon or epiphenomenon? Cells 2019, 8, 896.
- Mulkearns-Hubert, E.E.; Reizes, O.; Lathia, J.D. Connexins in cancer: Jekyll or Hyde? Biomolecules 2020, 10, 1654.
- Trosko, J.E.; Chang, C.-C.; Upham, B.L.; Tai, M.-H. Ignored hallmarks of carcinogenesis: Stem cells and cell-cell communication. Ann. N. Y. Acad. Sci. 2004, 1028, 192–201.
- Aasen, T.; Mesnil, M.; Naus, C.C.; Lampe, P.D.; Laird, D.W. Gap junctions and cancer: Communicating for 50 years. Nat. Rev. Cancer 2016, 16, 775–788.
- Axelsen, L.N.; Calloe, K.; Holstein-Rathlou, N.-H.; Nielsen, M.S. Managing the complexity of communication: Regulation of gap junctions by post-translational modification. Front. Pharmacol. 2013, 4, 130.
- Babica, P. Gap Junctional Intercellular Communication: In Vitro Assessment of Hazardous and Beneficial Effects of Chemical. Docent Habilitation Thesis, Masaryk University, Brno, Czech Republic, 2017. Available online: https://is.muni.cz/do/rect/habilitace/1431/12437/habilitacni_prace/Babica_2017_FINAL_TEXT_.pdf (accessed on 16 June 2021).
- Trosko, J.E. The gap junction as a “Biological Rosetta Stone”: Implications of evolution, stem cells to homeostatic regulation of health and disease in the Barker hypothesis. J. Cell Commun. Signal. 2011, 5, 53–66.
- Trosko, J.E. Gap junctional intercellular communication as a biological “Rosetta stone” in understanding, in a systems biological manner, stem cell behavior, mechanisms of epigenetic toxicology, chemoprevention and chemotherapy. J. Membr. Biol. 2007, 218, 93–100.
- Upham, B.L.; Trosko, J.E. Oxidative-dependent integration of signal transduction with intercellular gap junctional communication in the control of gene expression. Antioxid. Redox Signal. 2009, 11, 297–307.
- Abbaci, M.; Barberi-Heyob, M.; Blondel, W.; Guillemin, F.; Didelon, J. Advantages and limitations of commonly used methods to assay the molecular permeability of gap junctional intercellular communication. Biotechniques 2008, 45, 33–62.
- Maes, M.; Yanguas, S.C.; Willebrords, J.; Vinken, M. Models and methods for in vitro testing of hepatic gap junctional communication. Toxicol. In Vitro 2015, 30, 569–577.
- Iyyathurai, J.; Himpens, B.; Bultynck, G.; D’hondt, C. Calcium wave propagation triggered by local mechanical stimulation as a method for studying gap junctions and hemichannels. In Gap Junction Protocols; Methods in Molecular Biology; Vinken, M., Johnstone, S.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 1437, pp. 203–211. ISBN 978-1-4939-3664-9.
- El-Fouly, M.H.; Trosko, J.E.; Chang, C.-C. Scrape-loading and dye transfer: A rapid and simple technique to study gap junctional intercellular communication. Exp. Cell Res. 1987, 168, 422–430.
- Upham, B.L.; Sovadinova, I.; Babica, P. Gap junctional intercellular communication: A functional biomarker to assess adverse effects of toxicants and toxins, and health benefits of natural products. J. Vis. Exp. 2016, 2016, e54281.
- Babica, P.; Sovadinová, I.; Upham, B.L. Scrape loading/dye transfer assay. In Gap Junction Protocols; Vinken, M., Johnstone, S.R., Eds.; Springer: New York, NY, USA, 2016; Volume 1437, pp. 133–144. ISBN 978-1-4939-3664-9.
- Dydowiczová, A.; Brózman, O.; Babica, P.; Sovadinová, I. Improved multiparametric scrape loading-dye transfer assay for a simultaneous high-throughput analysis of gap junctional intercellular communication, cell density and viability. Sci. Rep. 2020, 10, 730.
- Upham, B.L.; Park, J.-S.; Babica, P.; Sovadinova, I.; Rummel, A.M.; Trosko, J.E.; Hirose, A.; Hasegawa, R.; Kanno, J.; Sai, K. Structure-activity-dependent regulation of cell communication by perfluorinated fatty acids using in vivo and in vitro model systems. Environ. Health Perspect. 2009, 117, 545–551.
- Tsao, M.S.; Smith, J.D.; Nelson, K.G.; Grisham, J.W. A diploid epithelial cell line from normal adult rat liver with phenotypic properties of “oval” cells. Exp. Cell Res. 1984, 154, 38–52.
- Shafritz, D.A.; Dabeva, M.D. Liver stem cells and model systems for liver repopulation. J. Hepatol. 2002, 36, 552–564.
- De Feijter, A.W.; Ray, J.S.; Weghorst, C.M.; Klaunig, J.E.; Goodman, J.I.; Chang, C.C.; Ruch, R.J.; Trosko, J.E. Infection of rat liver epithelial cells with v-Ha-ras: Correlation between oncogene expression, gap junctional communication, and tumorigenicity. Mol. Carcinog. 1990, 3, 54–67.
- Coleman, W.B.; McCullough, K.D.; Esch, G.L.; Faris, R.A.; Hixson, D.C.; Smith, G.J.; Grisham, J.W. Evaluation of the differentiation potential of WB-F344 rat liver epithelial stem-like cells in vivo. Differentiation to hepatocytes after transplantation into dipeptidylpeptidase-IV-deficient rat liver. Am. J. Pathol. 1997, 151, 353–359.
- Grisham, J.W.; Coleman, W.B.; Smith, G.J. Isolation, culture, and transplantation of rat hepatocytic precursor (stem-like) cells. Proc. Soc. Exp. Biol. Med. 1993, 204, 270–279.
- Malouf, N.N.; Coleman, W.B.; Grisham, J.W.; Lininger, R.A.; Madden, V.J.; Sproul, M.; Anderson, P.A. Adult-derived stem cells from the liver become myocytes in the heart in vivo. Am. J. Pathol. 2001, 158, 1929–1935.