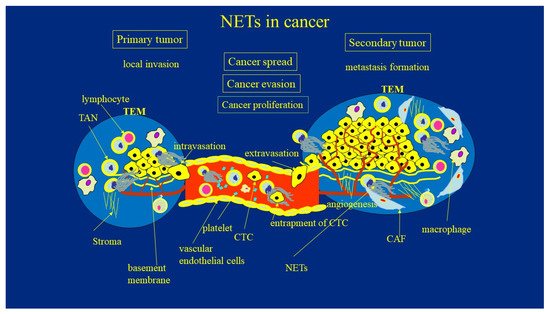
Tumor cells can migrate and intravasate the blood or lymph vasculature. They can survive within the circulation, then extravasate at distant sites. The factors determining adhesion strength, which might influence the ability of cells to transmigrate through an endothelial cell monolayer and the basement membrane, are poorly understood. Recent studies have highlighted that these processes are driven not only by signals from cancer cells, but are also modified by signals from components of the TME [
55]. Current evidence suggests that NETs may play a crucial role in the hematogenous spread of tumors. Jung et al. [
41] showed that NETs promoted tumor growth, metastasis, and angiogenesis of the pancreatic cancer cell line (AsPC-1). NETs used as chemoattractans stimulated AsPC-1 cell migration (in a Matrigel-coated invasion chamber) better then intact neutrophils. These effects were abrogated by histone-binding agents (heparin, polysialic acid), DNAse I, and Toll-like receptor neutralizing antibodies. Antibodies against both TLR2 and TLR4 significantly inhibited NET-mediated AsPC-1 cell migration. Although not unexpectedly, these results support the opinion that TLR2 and TLR4 participate in tumor transmigration. In patients with pancreatobiliary malignancy, elevated NET markers correlated with hypercoagulability makers. Histone–DNA complexes were used as markers of NETs. Another component of NETs, histones, significantly increased the endothelial cell proliferation and the formation of new blood vessels in a dose-dependent manner. Application of histone-binding agents abrogated histone-induced angiogenesis [
41]. The same directionality of the effect was observed by Tohme et al., who reported that the chemotactic factor released during NET formation may stimulate proliferation and migration of cancer cells [
36]. Finally, the transmigration mechanisms were explained by Kołaczkowska et al., who observed the adherence of circulating NETs to blood, resulting in increased cancer extravasation efficiency, which would enable cancer cells to cross the endothelial barrier [
61]. On the other hand, the previously mentioned report of Park et al. suggested that not DNA itself, but rather NET-related proteases are responsible for this effect [
59]. Such a discrepancy may be explained by the fact that such a structure as complex as the one between NETs and the locally concentrated enzymes, must be taken as an inseparable assembly, rather than a conglomerate of individual components. Once in the circulation, tumor cells become entrapped by NETs DNA threads. Through the use of cecal ligation, Cools-Lartigue et al. [
62] demonstrated the presence of circulating lung carcinoma cells wrapped in NET DNA conglomerates in a murine model of infection. Consequently, circulating “packages” were seeded in the liver, forming micrometastases within 48 h and secondary liver cancer 2 weeks after the cancer cell injection. DNAse or NE inhibitors abrogated the effects [
62]. Evidence consistent with these observations was provided by Najmeh et al. from the same group, who found a significant association between upregulation of β1-integrin and NET-related entrapment of circulating lung carcinoma cells, further facilitating metastasis formation and cancer spread [
63]. Whatever the precise basis of this mechanism is, it appears that inflammatory mediators harbored by neutrophils may be responsible for insufficient clearance of circulating cells [
64]. NETs’ entrapping abilities can be, at least partially, attributed to the ability to adhere to DNA mesh carried by the variety of integrins expressed on the surface of cancer cells. Such interaction was completely abrogated by DNase 1 [
65]. Furthermore, the TAN-CTC adhesion process facilitates cancer cell extravasation through the breaking of the transendothelial barrier [
66]. The proposed adhesive interaction between circulating neoplastic cells and TANs leads to the increased endothelial cell contraction, permeability, and malignant cell extravasation [
66]. A multi-level model shed new light on the fundamental processes elucidating the role of NETs in cancer invasions, transport, and transendothelial migration, thus taking into account specific NET–cell adhesion, ECM–tumor–NET interaction, and intracellular signaling [
67,
68,
69,
70,
71]. Further studies, however, are still warranted to explore these issues.