Hydrogen sulfide (H2S) is predominantly considered as a gaseous transmitter or signaling molecule in plants.
- abiotic stress
- hydrogen sulfide
1. Introduction
Hydrogen sulfide has been known as a crucial player during various plant cellular and physiological processes and has been gaining unprecedented attention from researchers since decades. They regulate growth and plethora of plant developmental processes such as germination, senescence, defense, and maturation in plants. Owing to its gaseous state, they are effectively diffused towards different parts of the cell to counterbalance the antioxidant pools as well as providing sulfur to cells. H2S participates actively during abiotic stresses and enhances plant tolerance towards adverse conditions by regulation of the antioxidative defense system, oxidative stress signaling, metal transport, Na+/K+ homeostasis, etc. They also maintain H2S-Cys-cycle during abiotic stressed conditions followed by post-translational modifications of cysteine residues. Besides their role during abiotic stresses, crosstalk of H2S with other biomolecules such as NO and phytohormones (abscisic acid, salicylic acid, melatonin, ethylene, etc.) have also been explored in plant signaling.
2. H2S-Mediated Mechanism of Action in Plants
2.1. Role of H2S in Post-Translational Modification of Cysteine Residues and Protein Sulfidation
2.1.1. Protein Persulfidation
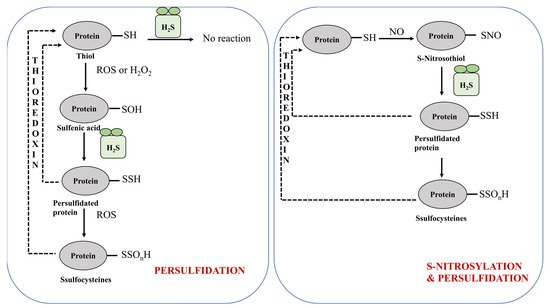
2.1.2. Protein Persulfidation in Plants
3. H2S-Signaling during Abiotic Stresses
3.1. H2S-Signaling during Heavy Metal Stresses
3.2. H2S-Signaling during Salinity Stress
3.3. H2S-Signaling during Drought/Osmotic Stress
3.4. H2S-Signaling during Temperature Stress
3.5. H2S-Signaling during Nutritional Stress
This entry is adapted from the peer-reviewed paper 10.3390/hydrogen2030017