Fucoidans are cell wall polysaccharides found in various species of brown seaweeds. They are fucose-containing sulfated polysaccharides (FCSPs) and comprise 5–20% of the algal dry weight. Fucoidans possess multiple bioactivities, including antioxidant, anticoagulant, antithrombotic, anti-inflammatory, antiviral, anti-lipidemic, anti-metastatic, anti-diabetic and anti-cancer effects. Dietary fucoidans provide small but constant amounts of FCSPs to the intestinal tract, which can reorganize the composition of commensal microbiota altered by FCSPs, and consequently control inflammation symptoms in the intestine. Although the bioactivities of fucoidans have been well described, there is limited evidence to implicate their effect on gut microbiota and bowel health.
- fucoidan
- intestinal function
- gut microbiota
- inflammation
- immunity
1. Introduction
“Fucoidans” refer to fucose-containing polysaccharides with sulfate groups, extracted from brown seaweed species such as Fucus vesiculosus, Cladosiphon okamuranus, Laminaria japonica and Undaria pinnatifida [1]. It is well-known that fucoidans are present in the seaweed cell wall to sustain cell membrane stability and protect their structure against dehydration [2]. The structure of fucoidans has been correlated with their biological and immunological activities, such as antioxidant, anti-coagulant, anti-thrombotic, anti-inflammatory, anti-viral, anti-lipidemic, anti-metastatic, anti-diabetic and anti-cancer effects [3]. Although fucoidan extracts have been widely used in various health care products including food supplements [4] and cosmetics [5], their bioactivities were reported to be sensitive to alterations in their structural composition. Moreover, understanding their functional properties are complicated since it is dependent on fucoidan extraction methods and the different seaweed species [6]. Some studies have demonstrated that functional characteristics of fucoidan extracts are closely linked to their structural formula, compositions of sugars, sulfate content and sulfate group positioning [3][7]. Therefore, more knowledge about structural and chemical characteristics of fucoidan is essential to better understand functional properties of the fucoidan.
2. Influence of Fucoidans on Intestinal Bowel Function
2.1. Effect on “Non-Immune Cells”
The intestinal epithelial barrier includes various types of cells, such as enterocytes, goblet cells, Paneth cells, and enteroendocrine cells, which are derived from multipotent intestinal stem cells (ISCs). A recent study showed that dietary components can also affect the differentiation of intestinal progenitor cells [8]. Lgr5+ ISCs can only recognize digested fructose and further differentiate into absorptive (i.e., enterocytes) or secretory progenitors (i.e., tuft cells, goblet cells, and Paneth cells), respectively. In addition, the intake of carbohydrates and proteins ameliorates intestinal epithelial injury by increasing membrane permeability [9]. Enterocytes line the surface of the epithelium in the small and large intestines, joined to each other by tight junctions. They are known to interact with nutrients including polysaccharides through antigen uptake and endocytosis [10]. Another study demonstrated that fucoidan extracted from Sargassum cinereum reduced the growth of the colon cancer cell line Caco-2, which has characteristics similar to enterocytes with a brush border epithelial layer [11]. In a study using human enterocyte-like HT-29-luc cells treated with extracts from Undaria pinnatifid, higher superoxide anion radical scavenging capacities and increased cell viability were shown compared to control groups [12]. Other studies on Caco-2 cell lines [13] showed that the eggshell membrane protein from chitosan/fucoidan nanoparticles reduced NO products and expression levels of TNF-α and IL-6, as well as having increased the paracellular permeability of fluorescein isothiocyanate-dextran in IECs, suggesting the importance of increasing bioavailability of fucoidan extracts to optimize their efficacy on immune response.
2.2. Effect on “Immune Cells”
Fucoidan has also been reported to modulate immune cell counts and their functions. A study employed immunohistochemistry to identify fucoidan-positive cells and their cell types in the small intestine of rats fed 2% fucoidan. The authors detected fucoidan and ED1 (macrophage marker) double-positive cells, indicating that intestinal macrophages may be the main cell type to internalize fucoidan [14]. A recent study showed that fucoidan treatment also affects the production of intracellular reactive oxygen species and recruitment of macrophages and neutrophils in LPS-induced RAW 264.7 murine macrophage cell lines and zebrafish larvae [15]. In the case of dendritic cells (DCs), significantly lower expressions of MHCII and CD86 was observed in bone marrow-derived DCs of non-obese diabetic mice fed fucoidan compared to those in the control group [16].
3. Fucoidan Structure
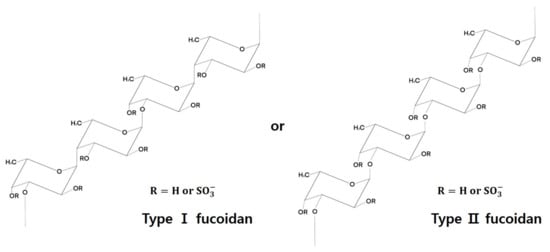
Although there is considerable variation among different algal species in terms of structural and chemical composition of fucoidans, they generally comprise the (1→3)-linked α-L-fucopyranosyl backbone structure, and occasionally both (1→3)-linked and (1→4)-linked α-L-fucopyranosyl structures [6] (Figure 1).
4. Functional Effects of Fucoidans
4.1. As an “Energy Sources”
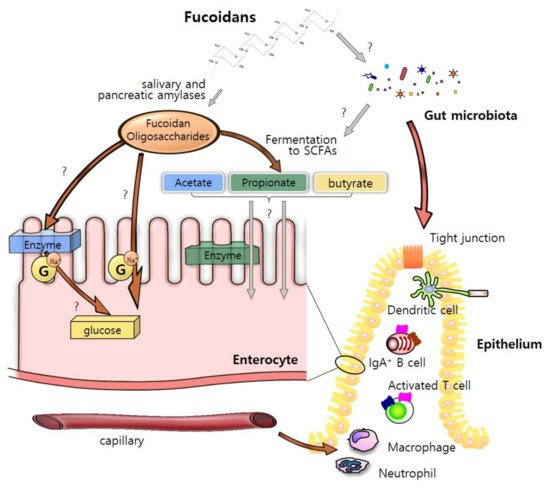
Fucoidan as a bioactive compound with high molecular weight and sulfated polysaccharides [18]. Fucoidan extracts contain many sugars including fucose, galactose, xylose, mannose, and glucose [3]. While fucose is generally rich in most brown seaweed species, the proportions of the saccharides vary among different species [19] (Table 2). Microvilli and plasma membranes of enterocytes play a critical role in the digestion and absorption of fucoidan [14]. There might be three possible routes of fucoidan oligosaccharides through intestinal epithelium. Many digestive enzymes including amylase, protease and lipase are attached to the plasma membrane and help in breakdown of fucoidans to the constituent monosaccharides [20][21]. These compounds may infiltrate enterocytes by active transport via Na+-dependent glucose transporters [22]. In addition, they could be directly recognized and transported into the epithelium by endocytosis of IECs [23]. It is also assumed that they are absorbed as short-chain fatty acids (SCFAs) by the fermentation by-products from fucoidan [24]. The transported nutrients from fucoidan as energy sources could enhance intestinal barrier integrity and maintain intestinal homeostasis by interacting with various intestinal cells (Figure 2).
4.2. As an “Immune Regulators”
Dietary fucoidans have been reported to have enhanced bioactivity and play pivotal roles in improving the gut health of animals, including humans [25], mice [26], livestock [27], and fish [28] (Table 1).
| Species | Fucoidan Sources | Dose | Tissues | Results | References |
|---|---|---|---|---|---|
| Human | Fucus vesiculosus and Undaria pinnatifida |
1 g/d | feces | increase in fecal lysozyme | [25] |
| C57BL/6 mice | Fucus vesiculosus | 5 mg/mL | colon, spleen, and feces | reduced diarrhea and fecal blood loss lower in colon and spleen weight decreases in IL-1α, IL-1β, IL-10, MIP-1α, MIP-1β, G-CSF and GM-CSF in the colon tissue |
[26] |
| Newly weaned pig | Laminaria spp. | 240 ppm | colon | increased intestinal villous height and the ratio of villus height to crypt depth | [27] |
| Nile tilapia | Fucus vesiculosus | 0.1% 0.2%, 0.4%, or 0.8% in basal diet | intestine | improved WG and SGR increases in organosomatic index in the intestine higher IEL and IEC counts |
[28] |
| Newly weaned pig | Laminaria spp. | 2.8 g/kg | feces | increased the coefficient of total tract apparent digestibility decreased counts of Escherichia coli |
[29] |
| C57BL/6J mice | Ascophyllum nodosum | 400 mg/kg | colon | decreases in TNF-α, IL-1β, IL-6, and IL-10 | [30] |
| C57BL/6 mice | Fucus vesiculosus | 400 mg/kg | Colon and feces | reduction in diarrhea and fecal blood decreased the infiltration of inflammatory cells and the expression levels of TNF-α and IL-1β in the colon tissue |
[26] |
5. “Fucoidan-Microbiota-Intestine” Axis
5.1. Steady-State Condition
Fucoidan extracted from diverse brown seaweed species can beneficially influence host intestinal conditions by mediating the changes in the composition of commensal microbiota. Undaria pinnatifida (wakame) enriches Bifidobacterium longum, a well-known prebiotic that affects host metabolic disorders by enhancing glucagon-like peptide 1 (GLP-1) absorption in the intestine [31][32]. Mice fed with fucoidan from Ascophyllum nodosum showed a high proportion of Lactobacillus species, which modulate several host intestinal immunity-related processes, such as intestinal epithelial cell regeneration [33][34].
5.2. Disease Condition
Inflammatory bowel diseases (IBD), including ulcerative colitis (UC) and Crohn’s disease (CD), are the most common intestinal disorders. In particular, IBD is a disease with an increasing worldwide incidence rate, and is characterized by multiple inflammatory reactions of unknown cause in the intestine. Two different fucoidan preforms, the fucoidan-polyphenol complex and depyrogenated fucoidan (DPF), have been shown to ameliorate DSS-induced acute colitis through the downregulation of pro-inflammatory cytokines, leading to decreased pathohistological scores [26]. Fucoidan extracts from Chnoospora minima also improve host inflammation symptoms by inhibiting the nitrous oxide (NO)-mediated expression of prostaglandin (PG) E2 in zebrafish [35]. Another intestinal disease that seriously threatens human health is colorectal cancer, which is the third most common cancer worldwide. Emerging evidence suggests that fucoidan may be a prospective anti-cancer agent. Although the efficacy of fucoidan uptake in intestinal epithelial cells varies based on its molecular size, fucoidan extracted from Cladosiphon okamuranus has been shown to enhance the survival rate in mice of a colorectal tumor-bearing model [36]. Fucoidan can control cell viability and the cellular cycle by downregulating the insulin-like growth factor (IRF)-I receptor via the Ras/Raf/ERK pathway [37]. Fucoidan extracted from Sargassum cinereum enhances reactive oxygen species (ROS) production and, consequently, inhibits cell proliferation by increasing the permeability of the mitochondrial membrane and the efficacy of apoptosis in Caco-2 cells [11].
6. Conclusions and Perspectives
This entry is adapted from the peer-reviewed paper 10.3390/md19080436
References
- Fitton, J.H. Therapies from fucoidan; Multifunctional marine polymers. Mar. Drugs 2011, 9, 1731–1760.
- Vo, T.S.; Kim, S.K. Fucoidans as a natural bioactive ingredient for functional foods. J. Funct. Foods 2013, 5, 16–27.
- Li, B.; Lu, F.; Wei, X.J.; Zhao, R.X. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695.
- Ikeguchi, M.; Saito, H.; Miki, Y.; Kimura, T. Effect of fucoidan dietary supplement on the chemotherapy treatment of patients with unresectable advanced gastric cancer. J. Cancer Ther. 2015, 6, 7.
- Fitton, J.H.; Dell’Acqua, G.; Gardiner, V.-A.; Karpiniec, S.S.; Stringer, D.N.; Davis, E. Topical benefits of two fucoidan-rich extracts from marine macroalgae. Cosmetics 2015, 2, 66–81.
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs 2011, 9, 2106–2130.
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. Isolation of fucoidan from Sargassum polycystum brown algae: Structural characterization, in vitro antioxidant and anticancer activity. Int. J. Biol. Macromol. 2017, 102, 405–412.
- Kishida, K.; Pearce, S.C.; Yu, S.; Gao, N.; Ferraris, R.P. Nutrient sensing by absorptive and secretory progenies of small intestinal stem cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G592–G605.
- Snipe, R.M.J.; Khoo, A.; Kitic, C.M.; Gibson, P.R.; Costa, R.J.S. Carbohydrate and protein intake during exertional heat stress ameliorates intestinal epithelial injury and small intestine permeability. Appl. Physiol. Nutr. Metab. 2017, 42, 1283–1292.
- Snoeck, V.; Goddeeris, B.; Cox, E. The role of enterocytes in the intestinal barrier function and antigen uptake. Microbes Infect. 2005, 7, 997–1004.
- Narayani, S.S.; Saravanan, S.; Ravindran, J.; Ramasamy, M.S.; Chitra, J. In vitro anticancer activity of fucoidan extracted from Sargassum cinereum against Caco-2 cells. Int. J. Biol. Macromol. 2019, 138, 618–628.
- Nemoto, M.; Kuda, T.; Eda, M.; Yamakawa, H.; Takahashi, H.; Kimura, B. Protective effects of mekabu aqueous solution fermented by Lactobacillus plantarum Sanriku-SU7 on human enterocyte-like HT-29-luc cells and DSS-induced murine IBD model. Probiotics Antimicrob. Proteins 2017, 9, 48–55.
- Lee, M.-C.; Huang, Y.-C. Soluble eggshell membrane protein-loaded chitosan/fucoidan nanoparticles for treatment of defective intestinal epithelial cells. Int. J. Biol. Macromol. 2019, 131, 949–958.
- Nagamine, T.; Nakazato, K.; Tomioka, S.; Iha, M.; Nakajima, K. Intestinal absorption of fucoidan extracted from the brown seaweed, Cladosiphon okamuranus. Mar. Drugs 2015, 13, 48–64.
- Jeong, J.-W.; Hwang, S.J.; Han, M.H.; Lee, D.-S.; Yoo, J.S.; Choi, I.-W.; Cha, H.-J.; Kim, S.; Kim, H.-S.; Kim, G.-Y.; et al. Fucoidan inhibits lipopolysaccharide-induced inflammatory responses in RAW 264.7 macrophages and zebrafish larvae. Mol. Cell. Toxicol. 2017, 13, 405–417.
- Xue, M.; Liang, H.; Ji, X.; Liu, Y.; Ge, Y.; Hou, L.; Sun, T. Fucoidan prevent murine autoimmune diabetes via suppression TLR4-signaling pathways, regulation DC/Treg induced immune tolerance and improving gut microecology. Nutr. Metab. 2019, 16, 87.
- Shang, Q. Revisit the effects of fucoidan on gut microbiota in health and disease: What do we know and what do we need to know? Bioact. Carb. Diet. Fibre 2020, 23, 100221.
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from Fucoidan: An Update. Mar. Drugs 2015, 13, 5920–5946.
- Wu, L.; Sun, J.; Su, X.T.; Yu, Q.L.; Yu, Q.Y.; Zhang, P. A review about the development of fucoidan in antitumor activity: Progress and challenges. Carbohydr. Polym. 2016, 154, 96–111.
- Sajina, K.A.; Sahu, N.P.; Varghese, T.; Jain, K.K. Fucoidan-rich Sargassum wightii extract supplemented with α-amylase improve growth and immune responses of Labeo rohita (Hamilton, 1822) fingerlings. J. Appl. Phycol. 2019, 31, 2469–2480.
- Zhang, C.; Kim, S.-K. Research and application of marine microbial enzymes: Status and prospects. Mar. Drugs 2010, 8, 1920–1934.
- Mabate, B.; Daub, C.D.; Malgas, S.; Edkins, A.L.; Pletschke, B.I. Fucoidan structure and Its impact on glucose metabolism: Implications for diabetes and cancer therapy. Mar. Drugs 2021, 19, 30.
- Chen, C.-H.; Lin, Y.-S.; Wu, S.-J.; Mi, F.-L. Mutlifunctional nanoparticles prepared from arginine-modified chitosan and thiolated fucoidan for oral delivery of hydrophobic and hydrophilic drugs. Carbohydr. Polym. 2018, 193, 163–172.
- Sun, T.; Xue, M.; Yang, J.; Pei, Z.; Zhang, N.; Qin, K.; Liang, H. Metabolic regulation mechanism of fucoidan via intestinal microecology in diseases. J. Sci. Food Agric. 2021, 101, 4456–4463.
- Cox, A.J.; Cripps, A.W.; Taylor, P.A.; Fitton, J.H.; West, N.P. Fucoidan supplementation restores fecal lysozyme concentrations in high-performance athletes: A pilot study. Mar. Drugs 2020, 18, 412.
- Lean, Q.Y.; Eri, R.D.; Fitton, J.H.; Patel, R.P.; Gueven, N. Fucoidan extracts ameliorate acute colitis. PLoS ONE 2015, 10, e0128453.
- Walsh, A.M.; Sweeney, T.; O’Shea, C.J.; Doyle, D.N.; O’Doherty, J.V. Effect of dietary laminarin and fucoidan on selected microbiota, intestinal morphology and immune status of the newly weaned pig. Br. J. Nutr. 2013, 110, 1630–1638.
- Mahgoub, H.A.; El-Adl, M.A.M.; Ghanem, H.M.; Martyniuk, C.J. The effect of fucoidan or potassium permanganate on growth performance, intestinal pathology, and antioxidant status in Nile tilapia (Oreochromis niloticus). Fish Physiol. Biochem. 2020, 46, 2109–2131.
- O’Doherty, J.V.; Dillon, S.; Figat, S.; Callan, J.J.; Sweeney, T. The effects of lactose inclusion and seaweed extract derived from Laminaria spp. on performance, digestibility of diet components and microbial populations in newly weaned pigs. Anim. Feed Sci. Technol. 2010, 157, 173–180.
- Wang, L.; Ai, C.; Wen, C.; Qin, Y.; Liu, Z.; Wang, L.; Gong, Y.; Su, C.; Wang, Z.; Song, S. Fucoidan isolated from Ascophyllum nodosum alleviates gut microbiota dysbiosis and colonic inflammation in antibiotic-treated mice. Food Funct. 2020, 11, 5595–5606.
- Yoshinaga, K.; Maruya, R.; Koikeda, T.; Nakano, T. Effects of Undaria pinnatifida (wakame) on the human intestinal environment. Funct. Foods Health Dis. 2018, 8, 488–504.
- Wei, P.; Yang, Y.; Li, T.; Ding, Q.; Sun, H. A engineered Bifidobacterium longum secreting a bioative penetratin-glucagon-like peptide 1 fusion protein enhances glucagon-like peptide 1 absorption in the intestine. J. Microbiol. Biotechnol. 2014, 24, 10.
- Shang, Q.; Shan, X.; Cai, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan modulates the gut microbiota in mice by increasing the abundance of Lactobacillus and Ruminococcaceae. Food Funct. 2016, 7, 3224–3232.
- Lee, Y.S.; Kim, T.Y.; Kim, Y.; Lee, S.H.; Kim, S.; Kang, S.W.; Yang, J.Y.; Baek, I.J.; Sung, Y.H.; Park, Y.Y.; et al. Microbiota-derived lactate accelerates intestinal stem-cell-mediated epithelial development. Cell Host Microbe 2018, 24, 833–846.e6.
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Samarakoon, K.W.; Lee, W.W.; Kim, H.S.; Kang, N.; Ranasinghe, P.; Lee, H.S.; Jeon, Y.J. A fucoidan fraction purified from Chnoospora minima; a potential inhibitor of LPS-induced inflammatory responses. Int. J. Biol. Macromol. 2017, 104 Pt A, 1185–1193.
- Kim, E.J.; Park, S.Y.; Lee, J.Y.; Park, J.H. Fucoidan present in brown algae induces apoptosis of human colon cancer cells. BMC Gastroenterol. 2010, 10, 96.
- Kim, I.H.; Nam, T.J. Fucoidan downregulates insulin-like growth factor-I receptor levels in HT-29 human colon cancer cells. Oncol. Rep. 2018, 39, 1516–1522.