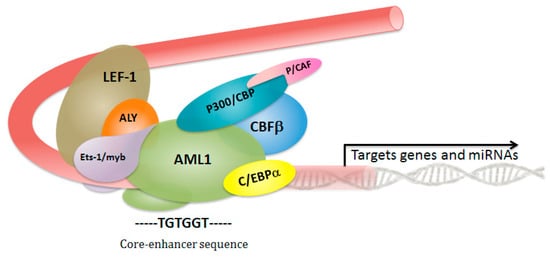
Current treatment guidelines for CBFL with t(8;21) do not take into account heterogeneity in these patients, and thus, all CBFL patients generally receive the same induction and consolidation treatments. Many comprehensive genetic analyses recognize that combination of several genetic alterations is associated with the development of CBFL, and is necessary for a better risk stratification in this leukemia. Although the spectrum of mutations for both CBFL subtypes is similar to the reported signature for AML [
96], gene expression and mutation profiling of CBFL identified t(8;21) and inv(16) patients as two distinct subgroups [
97], reflecting alternative signals activated in each subtype of CBFL [
98]. Moreover, 35% of CBFL patients have two or more mutations in tyrosine kinase (TK) genes coding for pathway effectors (especially
KIT,
FLT3, and
RAS genes); these findings highlight the multiclonality of CBFL.
NRAS is the most frequently mutated gene in CBFL, more common in
CBFB–MYH11 with a different spectrum of mutations, yet its mutations are not associated with outcome.
KIT mutations are found in ~40% of CBFL with t(8;21) and ~33% with inv(16); additionally, an enrichment of exon 17
KIT mutations has been documented in
RUNX1–RUNX1T1 patients, and are associated with worse outcome [
87,
99,
100,
101]. Recent large study created an “International CBF group index for t(8;21)” and validated a new risk scoring system, showing that older age, higher WBC index at diagnosis [
102], and KIT D816V/Y mutations were risk factors associated with treatment failure (relapse or death) [
103]. These studies strongly support the adverse effect of a
KIT mutation in the context of CBFL. In addition, a novel finding indicates that pseudodiploidy was also a risk factor in t(8;21). High-risk score patients may benefit from more intensive approaches in the first complete remission (CR1) [
103]. Mutations affecting
FLT3–ITD are present in only 3% of inv(16) AML, whereas they occur in 10% of t(8;21) leukemia patients. In addition to mutations in genes involving TK signaling, alterations in
MGA (MAX dimerization protein), a negative regulator of MYC signaling, were also recurrently identified in CBFL [
104]. Recent results identified CCND2 (cyclin D2) expression as key transmitter of
RUNX–RUNX1T1-driven AML, promoting cell cycle progression with the cooperation of the transcription factor Activator protein 1 (AP-1), and suggesting new potentially targetable complexes in CBFL [
14,
105]. Loss-of-function mutations in genes that regulate chromatin-modifying genes (
ASXL1/2,
EZH2,
KDM6A,
BCOR/BCORL1,
EED,
SETD2,
KMT2D,
KMT2C, and
CREBBP) or in genes implicated in the cohesin complex (
RAD21,
SMC1A,
SMC3,
STAG2) were observed almost exclusively in
RUNX1–RUNX1T1 AML. Cohesin mutations led to a state of increased chromatin accessibility of binding sites for master hematopoietic transcription factors such as AML1 [
106]. These findings suggest links between cohesin-mediated alterations in chromatin structure, or chromatin modifiers mutations, and cooperativity with the AML1–ETO fusion oncoprotein, even if cohesin mutations concerned less than 10% of CBFL [
15]. CBFL patients with mutations in the above members of the complex, responsible for sister chromatid cohesion during mitosis and DNA repair, lack evidence of aneuploidy or an increase rate of genetic instability without any effect on the outcome. Recently, mutations in
ASXL1 (additional sex combs like 1),
ASXL2 (additional sex combs like 2),
ZBTB7A (zinc finger and BTB domain conteining 7A),
CCND2, and
DHX15 (DEAH-box helicase 15) have been frequently identified in
RUNX1–RUNX1T1 but not in
CBFB–MYH11 AML patients [
14,
107].
ASXL1 or
ASXL2 truncating mutations, which inhibit myeloid differentiation and induce a myelodysplastic syndrome-like disease in mice [
108,
109], have been described in ~35% of t(8;21) while are absent in inv(16) AML [
15,
110,
111]. Of interest, chromatin modifier
ASXL1, as well as cohesin gene mutations, are co-occurring alterations significantly enriched in patients with mutated
RUNX1 AML [
112,
113]. The nature of cooperating mutations associated with t(8;21)-mediated leukemogenesis is evidenced by additional cytogenetic abnormalities such as trisomy 8 and 4, chromosome 9 deletion, and loss of one of the sex chromosomes [
114,
115,
116]. Increased dosage of the mutated
KIT (mapped at 4q12) can occur due to trisomy 4, leading to duplication of the mutant
KIT allele, and suggesting an additional contribution to leukemogenesis [
86]. These observations are supported by a higher dosage of N822K
KIT mutated allele linked to an increased segregation of minichromosomes derived from chromosome 4 that preserve the pericentromeric region containing the
KIT gene in the t(8;21) positive Kasumi-1 cell line [
117]. The most common additional cytogenetic features associated with t(8;21) include loss of either the X or Y chromosome in a disproportionally large number of cases (50–60%), and del(9)(q22) in 15–25% of patients. It has been proposed that haploinsufficiency must be occurring at genes located within shared sequences in the pseudoautosomal regions (PARs) on the X and Y chromosome. A critical event potentially explaining the high incidence of loss of sex chromosomes in t(8;21) may be the loss of
CSF2RA (colony stimulating factor 2 receptor alpha subunit) gene, encoding for the α subunit of the heterodimeric receptor CSF2 (colony-stimulating factor 2), which control granulopoiesis [
118]. However, given that the whole sex chromosome is typically missing and not only the individual
CSF2RA locus, it is likely that additional haploinsufficient factors on the sex chromosome are acting to enhance
RUNX1–RUNX1T1-associated leukemogenesis [
119]. Sex chromosome loss was reported as a favorable marker for two-year event-free survival (66.9% vs. 43.0%), and in another study showed a modestly favorable, but not significant, effect on disease-free survival (DFS) [
103]. Moreover, this last study found that patients with pseudodiploid karyotypes had worse outcome compared with those with hypodiploidy or hyperdiploidy [
103]. In contrast, loss of the Y chromosome showed shorter disease-free survival (DFS) for male patients [
120].