Global development has generated a plethora of unfavorable and adverse environmental factors for the living organisms in the ecosystem. Plants are sessile organisms, and they are crucial to sustain life on earth. Since plants are sessile, they face a great number of environmental challenges related to abiotic stresses, such as temperature fluctuation, drought, salinity, flood and metal contamination. Salinity and drought are considered major abiotic stresses that negatively affect the plants’ growth and production of useful content. However, plants have evolved various molecular mechanisms to increase their tolerance to these environmental stresses. There is a whole complex system of communication (cross-talk) through massive signaling cascades that are activated and modulated in response to salinity and drought. Secondary metabolites are believed to play significant roles in the plant’s response and resistance to salinity and drought stress. Until recently, attempts to unravel the biosynthetic pathways were limited mainly due to the inadequate plant genomics resources. However, recent advancements in generating high-throughput “omics” datasets, computational tools and functional genomics approach integration have aided in the elucidation of biosynthetic pathways of many plant bioactive metabolites.
Note: The following contents are extract from your paper. The entry will be online only after author check and submit it.
1. Introduction
Due to the plants’ sessility and inability to migrate in nature, they are more prone to being affected by a great number of abiotic stresses such as salinity, drought, heat and radiation. Those abiotic stresses are considered major challenges for the development and growth of the plants and potentially influence the yield and quality of the useful products that provide medicinal or nutritional benefits to humans [
1]. Hence, understanding the mechanisms by which plants react to abiotic stresses will aid in improving the quality and quantity of the rich and diverse natural products.
Many of these stressors induce elicitors that would eventually result in either negative or positive plant responses [
2]. For instance, a positive influence has been proved by Crisosto [
3] in a study on peaches; it showed that a higher fruit density was observed at lower levels of irrigation. On the contrary, abiotic stresses also have negative effects on the development and growth of the plant. For example, they can result in changing the reactive oxygen species (ROS) levels in the body and cause damage; these changes will force the plant to undergo programmed cell death (PCD) [
4]. Other examples of negative effects of abiotic stresses are the osmotic and ionic stresses that occur when plants are exposed to increased levels of salinity and water shortage [
5].
The plants tend to adapt to those abiotic stresses in order to survive and thrive [
6]. There are many players that contribute to the process of resistance, including signaling molecules, DNA-binding proteins and an enormous number of stress-inducible genes. Those biological players participate in the complex orchestration of the plants’ tolerance to stress at the molecular level. Stress-related molecules such as abscisic acid (ABA), jasmonic acid (JA) [
7], ROS, calcium ions, a variety of transcription factors (TFs) and other regulatory and functional proteins all interact together in a spectacular manner in order to perform a remarkable adaptation to the abiotic stresses [
8,
9]. This communication between secondary messengers, phytohormones, DNA-binding proteins and stress-inducible genes is known as “cross-talk” [
9,
10].
In response to abiotic stresses, such as salinity and drought, the genetic regulation also involves a complex organization of metabolic activities that involve the intracellular production of beneficial secondary metabolites. Those secondary metabolites facilitate the plant’s defense mechanism by conferring many biological activities such as protecting the plant against oxidative stress, and they are known as stress-protectant metabolites [
11,
12]. A number of studies have undisputedly determined that many secondary metabolites such as flavonoids, sesquiterpenes and phenolic acids have a substantial antioxidant activity role. Since the changing balance of the ROS in the plant occurs when plants are subjected to abiotic stress, these antioxidative secondary metabolites contribute to the defense mechanisms that the plant utilizes to overcome the harmful effects of stress [
13,
14,
15]. Many plant-based secondary metabolites provide beneficial uses as pharmaceuticals, agrochemicals, flavors, fragrances, colors, biopesticides and food additives [
16,
17]. There are many secondary metabolites with beneficial use as a dietary component or for medicinal purposes, including flavonoids (anthocyanins), alkaloids, terpenes and many others [
11,
12].
Throughout history, genetic manipulation techniques led to the improvement of the plant’s production and quality of useful compounds and resistance to abiotic stresses. Plant natural products have been considered essential phytochemicals that have been used extensively in the field of medicine throughout human history. Some of the well-known plant-extract-derived medicines are morphine, quinine, colchicine, codeine and many others [
18]. The anticancer drug that is known as Taxol is extracted from trees from the
Taxus genus [
12]. A remarkable number of studies on medicinal plants displayed crucial evidence that when they are subjected to abiotic stress, the accumulation of secondary metabolites is usually achieved [
19,
20,
21,
22]. As a result of these findings, the interest in plants’ defense mechanisms and their secondary metabolite production has expanded and been focused on by many researchers.
2. Generic Pathways for Plant Response to Abiotic Stresses
According to our current knowledge about stress signaling pathways, the generic signaling pathway for any given abiotic stress can be divided into the following major steps: signal perception, signal transduction, stress-responsive gene expression and the activation of physiological and metabolic responses [
23]. When plants are exposed to drought or salinity stresses, they produce signals that are perceived at the membrane level by ion channels, membrane receptors, receptor-like kinases and many other proteins. Upon perception, complex intracellular signals are then initiated and cause the generation of many messengers such as Ca
2+, reactive oxygen species (ROS), phytohormones and other stress signaling molecules [
24,
25,
26]. Those signals are then transduced further inside the cell and interact with other molecules that will ultimately lead to the induction of the stress-induced gene that encodes for proteins that will particularly enhance the plant’s tolerance to abiotic stresses directly or indirectly. The plant response to abiotic stresses can be described as the coordination of expression of genes that encode for a variety of products that contribute to the plant’s adaptation to abiotic stresses [
27,
28]. Those products could carry out any function that supports the plant in the adaptation to the stress; they could be, for instance, secondary metabolites that carry out essential protective functions against abiotic stresses, transcription factors that facilitate the activation of second-level stress-inducible genes or any other functional or regulatory molecules [
29].
Based on the protein products of the stress-inducible genes, they can be divided into two functional categories: (1) genes that encode for products that directly affect the plant’s tolerance to salinity and water deficit stresses, such as heat stress proteins (HSPs) or chaperones, LEA proteins, osmoprotectants, antifreeze proteins, detoxification enzymes and free-radical scavengers [
30], and (2) stress-responsive genes that encode for protein products that influence the abiotic stress resistance by regulating the expression of other downstream target genes and modulate the signal transduction cascades. The latter category includes genes that may produce phytohormones such as abscisic acid (ABA), jasmonic acid (JA), salicylic acid (SA) and ethylene [
7]. It also includes genes that encode for other signaling molecule products that contribute to the signaling cascade regulation of gene expression, such as the mitogen-activated protein kinases (MAPKs), calcium-dependent protein kinases (CDPKs) and a great number of stress-inducible transcription factors (
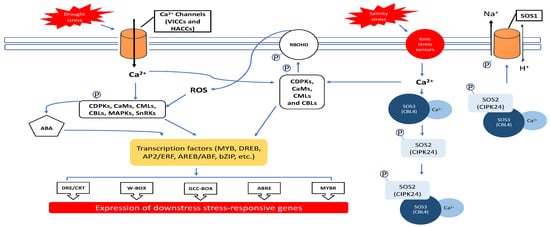
Figure 1).
Figure 1. Generic pathway for plant response to salinity and drought stresses in Arabidopsis thaliana cell. Salinity and drought elicit Ca2+ signals through the SOS pathway to achieve ionic homeostasis. When the ions are increased, it is first perceived by the ionic stress sensors, and then the Ca2+ levels are increased. The Ca2+ binds to the SOS3 protein and then interacts with the SOS2 protein kinase. The Ca2+–SOS3–SOS2 complex phosphorylates and activates the SOS1 membrane protein, which will cope with the ionic stress. This change in cytoplasmic Ca2+ level is sensed by calcium sensors (CDPKs, CaMs, CMLs and CBLs) that interact with their downstream signaling components, which could be RBOHD or transcription factors. The activation of RBOHD leads to the generation of ROS molecules, which in turn aid in the signaling mechanism through the interaction with many molecules such as phytohormones or several kinases. Those phytohormones (ABA) or the protein kinases interact with the stress-inducible TFs. Those stress-inducible TFs carry specific DNA-binding domains that bind to specific regulatory sequences found upstream of the stress-inducible gene and initiate transcription. Those genes will lead to several physiological and chemical changes that would enhance the plant’s tolerance to salinity and drought stresses.
2.1. Ca2+ and ROS Regulation during Salinity and Drought Exposure
The mechanism of genetic regulation in plants upon the exposure to drought or salinity as an abiotic factor involves the activation of several intricate signaling transduction pathways. There are many intracellular signaling molecules that are involved in the plant’s adaptation mechanism to salinity and drought stresses in which they interact together in a complex manner in order to produce a final protective role. Secondary messengers such as ROS and Ca
2+ are considered well-known and significant signaling molecules that are active at the first steps of the plant’s response to abiotic stresses [
31].
2.2. Ca2+ Ions
As reported by many articles, the most important universal secondary messenger is calcium ions (Ca
2+). As mentioned before, salinity and drought stresses cause ionic and osmotic stresses for the exposed plant. Hence, the plants adapt to the stresses by retaining the cellular homeostasis by dealing with osmotic and ionic stresses. It was discovered that the Salt Overly Sensitive (SOS) genes are a primer mediator for the ionic hemostasis in plants.
SOS3 gene encodes for a protein that acts as a receptor for cytosolic Ca
2+ [
32]. Increased levels of Ca
2+ lead to the activation of the cascade of events that would activate the genes responsible for protecting the plant from salt stress. For instance, the
SOS1 gene encodes for the plasma membrane Na
+/H
+ antiporter, and the
SOS1 gene is mediated by the
SOS2, which encodes for a calcium-dependent protein kinase (CDPK) [
32,
33,
34]. The plasma membrane Na
+/H
+ antiporter aids in protecting the plant from salinity stress by maintaining the osmotic balance by compartmentalizing Na
+ and chloride into vacuoles or extruding the excessive toxic Na
+ to the apoplast or surrounding tissues [
35,
36,
37,
38]. Along with SOS, Ca
2+ ions cooperate with other signaling molecules, including calcium-dependent protein kinases (CDPKs), which are considered very important phosphorylating agents in the plant response to abiotic stresses [
39].
CDPKs play important roles at the cellular and molecular level in plants: increasing the resistance and adaptation to abiotic stress and modulating metabolic pathways [
40]. The CDPKs play a significant role in the biosynthesis of important organic molecules by phosphorylating and activating a variety of biosynthetic metabolic pathways. Allwood discovered that a key enzyme in the defense response against abiotic stress, known as the phenylalanine ammonia lyase (PAL) enzyme, could be phosphorylated by a CDPK isoform [
41], indicating its connection to the secondary metabolite biosynthetic pathways and abiotic stress resistance.
2.3. Reactive Oxygen Species (ROS)
During abiotic stresses, such as salinity and drought, overproduction of ROS results in perturbation of the cellular redox state and thus poses the threat of developing the toxic event that is known as oxidative stress. However, ROS are usually present at a low level in many organelles, and they act as significant secondary messengers in the plant’s response to abiotic stresses and secondary metabolism. They are considered very effective secondary messengers due to their spatiotemporal flexibility [
31].
There is a synergistic effect in the plant’s defense system between Ca
2+ and ROS signaling [
42,
43]. The respiratory burst oxidase homolog (
RBOH) gene family encodes for an enzyme known as the nicotinamide adenine dinucleotide phosphate (NADPH) oxidase. This product enzyme is considered the major contributor of de novo synthesized ROS extracellularly. The
RBOH gene is regulated and activated by the previously mentioned calcium-dependent protein kinase (CDPK), which facilitates its phosphorylation. Thus, in order to produce more ROS, Ca
2+ levels ought to be increased. Vice versa, elevated levels of ROS lead to the increased concentration of Ca
2+ ions by the interaction of the H
2O
2 and hydroxyl radicals with the Ca
2+-permeable cation channels such as the voltage-independent Ca
2+-permeable cation channels (VICCs) and the hyperpolarization-activated Ca
2+-permeable cation channels (HACCs). This results in a noticeable transient increase in the cytosolic Ca
2+, which is involved in several fundamental physiological and regulatory activities [
26,
44,
45].
Similar to Ca
2+, ROS interact with other signaling molecules such as the MAPKs. The mitogen-activated protein kinase is one of many important signaling molecules that initiate cascades of events that are involved with transducing stress-related stimuli such as enhanced levels of ROS or calcium signaling. It was discovered that ROS upregulate the activity of the
ANP1 kinase in
Arabidopsis [
46]. and its functional homolog
NPK1 in tobacco [
43], which trigger the downstream MAPK cascades. Several molecular genetic experiments indicated that
ANP1 is responsible for the regulation and activation of
AtMPK3 and
AtMPK6 [
39]. The protein kinase products of MPK3/MPK6 are responsible for the direct phosphorylation of the transcription factors associated with the plant’s defense system, such as WRKY and ERF. Many other studies have proved the association between ROS and MAPKs in several plant models, indicating their function in the plant’s response to abiotic stresses [
47,
48].
3. Phytohormonal Regulation in Salinity and Water Deficit Stresses
Phytohormones, e.g., abscisic acid (ABA), jasmonic acid (JA), salicylic acid (SA) and ethylene [
7], play critical regulatory tasks in the plant’s response to abiotic stresses and metabolic pathways. Each hormone plays a variety of functions, and all together act in a concerted way rather than a stand-alone action. Generally, hormones are synthesized at low concentrations in the body of the plant, and they function as a controlling agent in different aspects of developmental and growth events throughout the plant’s cycle of life [
43].
3.1. Abscisic Acid
ABA contributes to a variety of regulatory networks involving plant’s response to drought and salinity stresses that cause osmotic and ionic stresses. Under conditions of osmotic stress, abscisic acid is generally considered as a stress signaling hormone, and the expression of stress-responsive genes in plants is primarily regulated by ABA-dependent and ABA-independent pathways based on the
cis-acting regulatory elements present in the promoter regions of the stress-responsive genes [
49,
50]. The
cis-acting elements are regulatory sequences found at the promoter region of many genes, and they are important because they act as physical targets for many transcription factors in order to activate the downstream target genes. The transcription factors usually carry a specific DNA-binding domain that binds to particular
cis-regulatory elements [
51]. In the ABA-dependent gene expression, the
cis-acting element is the ABA-responsive element (ABRE) and involves the function of the ABA-dependent ABRE-binding protein/ABRE-binding factor (AREB/ABF) transcription factors that target the sequence known as G-box-like cis-acting element with the sequence of ACGTGG/TC [
52].
Abscisic acid phytohormone regulates the activation of calcium channels that in turn lead to the activation of the previously mentioned enzyme NADPH oxidase, which has the ability to produce de novo ROS molecules. Moreover, ROS interacts with ABA and other hormones for the main purpose of defending the plant by regulating the signaling cascades and the expression of many stress-responsive genes. Such genes’ products are antioxidant enzymes with ROS termination functions such as
superoxide dismutase (SOD),
catalase (CAT),
peroxidase (POD) and
ascorbate peroxidase (APX) [
43].
3.2. Ethylene
Ethylene is a gaseous molecule and a well-studied plant phytohormone that carries out important molecular modulation responsible for enhancing the tolerance of the plant to abiotic stress and regulating several metabolic pathways for secondary metabolite biosynthesis. Under salinity and drought stresses, the accumulation of ethylene was observed in many plants. A considerable body of evidence suggests that ethylene functions by monitoring gene expression as a part of its signal transduction pathway. In the absence of ethylene, very important genes that contribute to the plant’s response to stress will not be expressed; on the contrary, they will be degraded by E3 ubiquitin ligase [
53,
54]. Those genes are known as
ethylene insensitive 2 (
EIN2) and
ethylene insensitive 3 (
EIN3); they are two key intracellular ethylene-mediated gene regulators in the plant’s defense response. With ethylene,
EIN2 accumulates and thus facilitates the accumulation of the EIN3 proteins. The EIN3 proteins have a very important role in the genetic activation of a crucial stress-inducible transcription factor known as ethylene response factor 1 (
ERF1) [
53,
55]. The
ERF1 then assists as a transcription factor and recognizes a GCC
cis-acting element in the promoter regions of ethylene-responsive genes and triggers transcription. This finding shows only one example of how ethylene as a plant hormone contributes to regulating gene expression in the plant’s response to abiotic stresses such as drought and salinity.
3.3. Jasmonic Acid and MeJA
Plant produce jasmonic acid (JA) as a plant hormone to regulate a variety of plant processes, including response to abiotic stresses and regulation of the biosynthetic pathways of many secondary metabolites [
12]. JA or JA-Ile signaling results in the activation of many TFs that regulate the expression of a great number of target genes through specific binding to particular cis-acting elements. Moreover, jasmonic acid and the methyl ester form of jasmonic acid (MeJA) have been proved to be able to act as an elicitor for the production of several secondary metabolites such as alkaloids, flavonoids, other phenolic compounds and many other compounds [
56].
4. Transcriptional Regulation during Salinity and Drought Stresses
Transcription factors are regulatory proteins that recognize particular regulatory sequences of bases located upstream of the coding region of the gene and thus control the transcription process. Every transcription factor is identified by conserved and specific DNA-binding domains that are specific for several
cis-acting elements. By binding to
cis-regulatory sequences, these proteins facilitate either the activation or repression of the transcription of downstream genes. With regards to transcriptional regulation of the transcription factors in response to abiotic stress, studies have found that many regulatory transcription factors play essential roles in multiple abiotic stress responses by regulating a large spectrum of downstream genes; hence, these genes are known as stress-inducible or -responsive genes [
57].
In the last few decades, considerable research has been conducted to identify and characterize various TFs involved in plant abiotic stress responses and metabolic pathways either in abscisic acid (ABA)-dependent pathways or ABA-independent pathways, such as AP2/EREBP, MYB, bHLH, WRKY, NAC and bZIP [
52,
58].
Arabidopsis thaliana utilizes over 5% of its genome to code for over 1500 TFs, roughly 45% of which are from families that are only specific to plants [
59]. In this review, detailed information regarding the transcriptional regulation of a variety of transcription factors, including the superfamilies AP2/EREBP, MYB and bHLH, is provided.
4.1. AP2/EREBP
The APETALA2/ethylene-responsive element binding protein (AP2/EREBP) family is a family of transcription factors thought to have a significant role in regulating gene expression at the transcription level [
23,
60,
61]. These TFs have a highly conserved DNA-binding domain known as AP2/ERF DNA-binding domain. This AP2/ERF (APETALA2/ethylene-responsive element binding factor) is a domain consisting of 50–70 amino acids and is found in many proteins in the plant kingdom, such as
A. thaliana ERF1, tobacco EREBPs (homolog of the ERF),
A. thaliana AP2,
A. thaliana C-repeat/dehydration-responsive element (DRE) binding factor 1 (CBF1 orDREB1) and DREB2 and
Arabidopsis thaliana and maize abscisic acid (ABA)-insensitive 4 (ABI4) proteins [
54,
61].
Upon stresses, the AP2/EREBP superfamily are induced through
cis-acting elements present at their promoter region. These regulatory elements include GCC box, ABRE, EBS, HSE, LRT and many other unknown binding sites that respond to a variety of abiotic stress stimuli such as AREBs, EIN3 and other transcription factors [
61,
62]. Thus, the transcriptional regulation pathways of this superfamily of transcription factor proteins are both ABA-dependent and -independent. For instance, promoter analysis studies on the DREB subfamily determined that it contains the most diverse
cis-acting elements that are related to abiotic stresses, including ABRE, MeJA response, TCA, ERE, MBS, HSE, TC-rich and LTR motifs. This finding suggests that those transcription factor-encoding genes are the most frequently expressed and involved in the signal transduction pathways related to abiotic stresses [
51,
54]. It also suggests that this subfamily is regulated through ABA-dependent pathways due to the presence of the ABRE
cis-acting element and through the ABA-independent pathway due to the availability of the other
cis-regulatory elements.
The DREB 2 subfamily of the AP2/EREBP superfamily can regulate the expression of multiple
responsive to dehydration (RD) genes by interacting with a GCC-like box called the DRE/CRT
cis-acting element found at the promoter region of many stress-inducible genes in a variety of plant species (e.g.,
RD29A,
RD17,
ERD10) [
63,
64,
65]. On the other hand, the ethylene-responsive factor (ERF) subfamily, which also belongs to the AP2/EREBP superfamily, activates an array of stress-responsive genes through the interaction with
cis-regulatory sequences that are found upstream of the coding region of many stress-inducible genes such as the ethylene-response element (ERE) or GCC box with the core sequence of AGCCGCC, jasmonic acid- and elicitor-responsive element (JERE), coupling element 1 (CE1) and CT-rich element [
6,
55,
66]. For instance, it was found that a certain ERF encoded by the
ERF74 binds to the RbohD promoter region and triggers its expression [
54,
67] and hence participates in the defense mechanism against salinity and drought by generating ROS that act as signaling molecules. There are many other pieces of evidence suggesting the solid regulatory effect of the AP2/ERF family of TFs in the plant response to abiotic stresses [
68].
4.2. MYB
The MYB family of transcription factors is one of the largest TF families in plants. A growing body of evidence has demonstrated the regulatory effects of the MYB family on many biological and biochemical activities such as defense and stress response regulation (Wu et al., 2019), primary and secondary metabolism regulation (Dubos et al., 2010; Wu et al., 2019) and the regulation of other plant-related developmental mechanisms. The MYB family of TFs is characterized by the presence of a conserved MYB DNA-binding domain.
A transcriptional regulatory network analysis of MYB TF family genes in rice was conducted by Smita et al [
69] in order to identify new links and characterize the interaction between the MYB family and the downstream target genes. In their study, they observed around 40 putative target genes for OsMYB that contain at least one MYB-binding region in their promoter. Out of the 40 target genes, 27 were found to have a high number of MYB-binding regions involved in drought-inducibility (MBS, CAACTG and TAACTG). This finding implies that the
OsMYB genes are strongly involved and have great regulatory roles in drought response at the transcriptional level [
69]. Studies have shown that
AtMYB2,
AtMYB74 and
AtMYB102 MYB genes are upregulated by drought stress [
70], which indicates that they play a role in the stress response regulation to protect the plant against the harmful effects of water deficit. Additionally, a study by Hasan et al. [
71] investigated the possibility of
MYB44 interacting directly with ABA signaling in cotton species. They concluded that the
MYB44 gene encodes for a TF that interacts with ABA receptor proteins such as PYLs, which bind to PP2Cs. Hence, the MYB44 has been determined to be a regulator in the ABA-dependent signaling pathway that is related to the activation and repression of the SNF1-related protein kinases 2 (SnRK2s) in which it phosphorylates several stress-inducible genes.
AtMYB20 enhances salt tolerance by repressing the expression of PP2Cs [
71,
72].
4.3. Basic Helix–Loop–Helix (bHLH)
The bHLH family of transcription factors is the second largest family in plants after the MYB family. In
Arabidopsis, 162 members of bHLH have been identified, and 167 have been identified in rice plants [
73]. As the name suggests, the bHLH family of transcription factors contains a highly conserved basic/helix–loop–helix unique structural domain consisting of two parts: basic amino acid and the helix–loop–helix region (HLH). Findings have determined that the bHLH protein predominantly attaches to the core DNA sequence motif called E-box (5-CANNTG 3), where N could be any nucleotide; the most common form of E-box is the G-box (CACGTG), which is also a binding sequence for the MYC family [
74,
75].
A growing body of evidence proves that the bHLH TFs are strictly involved in plants’ response to a variety of abiotic stresses such as drought and salinity [
74,
76]. Cui et al. [
77] conducted a transcriptome-wide expression analysis of bHLH genes from tea (
C. sinensis) and revealed that a total of 39
CsbHLH were upregulated under the exposure of the plant to drought stress. Moreover, transcriptomic studies in
Arabidopsis roots under salt exposure have shown that at least 15 bHLH genes are upregulated in response to the stress, which implicates their functional role in salt stress responses in plants. There is other evidence suggesting the crucial involvement of the bHLH transcription factor family in plants’ response to abiotic stresses.
This entry is adapted from the peer-reviewed paper 10.3390/app11156668