Chronic pain is a serious public health and socioeconomic problem that affects millions of people worldwide each year [
1]. Some chronic pain conditions can be a consequence of abnormal functioning of the nervous system. In particular, neuropathic pain arises from a lesion or disease of the somatosensory system at peripheral or central level [
2]. Peripheral neuropathic pain is the result of injuries to the peripheral nervous system induced by various conditions, including metabolic diseases, infections, mechanical trauma, and chemotherapy [
3]. Central neuropathic pain involves injuries to the central nervous system, commonly caused by spinal cord injury, stroke, and multiple sclerosis [
4]. The initial lesions involved in both forms of neuropathic pain trigger a cascade of changes that lead to maladaptive plasticity within the nervous system and inevitably to neuropathic hypersensitivity [
5]. The prevalence of chronic pain with neuropathic characteristics has been estimated to be in the range of 7–10% [
6] and it is expected to rise in the future [
3].
Commonly used analgesics for nociceptive pain, such as non-steroidal anti-inflammatory drugs (NSAIDs) and opioids, are less effective in neuropathic [
9] or cancer pain [
10]. In fact, among cancer patients, those with cancer-related neuropathic pain usually have more trouble to optimally control pain [
11]. First-line drug therapies for neuropathic pain include tricyclic antidepressants (TCAs), serotonin-norepinephrine reuptake inhibitors (SNRIs), gabapentinoids, and topical application of lidocaine or capsaicin [
12]. These medications can be effective, but carry the potential risk of many side effects [
12]. Opioids are lower-line drug options for neuropathic pain, as there is low evidence to support the use of opioids for long-term in chronic neuropathic pain, with possible benefit only for short-term use [
9,
13]. Therefore, despite the multiple agents available for the treatment of neuropathic pain, current treatment options provide an average reduction of neuropathic pain by 30–50% [
9]. The effectiveness of available drugs in the treatment of cancer pain is also very low [
14]. The difficulty in optimally treating these pain conditions has prompted significant research into new analgesic agents with fewer side effects.
Tetrodotoxin (TTX) is a potent non-peptide guanidinium neurotoxin that blocks voltage-gated sodium channels (VGSCs) found in puffer fish and other marine and terrestrial animals [
15]. VGSCs play an important role in pain and, in particular, numerous studies have shown that some of the TTX-sensitive VGSC subtypes are strongly implicated in the pathophysiology of chronic pain, especially neuropathic pain [
16]. Since TTX blocks this subset of VGSCs in a highly selective manner, this agent may have a potential role in relieving neuropathic pain. In fact, this agent is progressing in its preclinical and clinical development, having been evaluated in various clinical trials for the indications of cancer-related pain (NCT00725114, NCT00726011) and chemotherapy-induced neuropathic pain (NCT01655823).
2. Role of TTX-Sensitive Voltage-Gated Sodium Channels (VGSCs) in Neuropathic and Cancer Pain
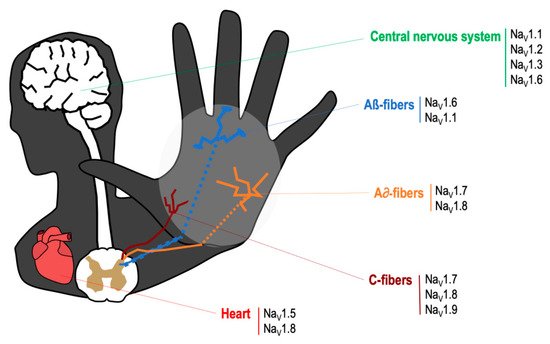
The VGSCs are integral membrane proteins composed of a 260 kDa α-subunit and one or more β-subunits, being the α-subunit responsible for forming the pore and for the main biophysical properties of the channel [
16]. Most VGSCs are located at peripheral and/or central nervous system (see
Figure 1), and are responsible for generating the Na
+ currents that lead to the initiation and propagation of neuronal action potentials [
17]. TTX binds to VGSCs by the interaction between the positively charged guanidine group in the TTX molecule with the negatively charged carboxylate residues that are placed around the outer vestibule of the channel; this binding occludes the channel pore and blocks Na
+ current [
18]. VGSCs are classified according to its sensitivity to TTX. Thus, TTX-sensitive VGSCs (Na
V1.1, Na
V1.2, Na
V1.3, Na
V1.4, Na
V1.6 and Na
V1.7) are blocked by nanomolar concentrations of this neurotoxin, whereas TTX-resistant VGSCs (Na
V1.5, Na
V1.8, Na
V1.9) are inhibited at micromolar concentrations [
16]. The mechanism of action by which TTX exerts its analgesic effect is by binding to the α-subunit within the outer vestibule of the VGSC, blocking the entry of Na
+ ions through the channel [
16,
18]. In this manner, TTX reduces the Na
+ ionic fluxes required for the initiation and conduction of nerve impulses.
Figure 1. Main location of voltage specific sodium channels in the nervous system.
Nociceptive pain has a physiological protective role in preventing tissue injury. Under normal conditions, primary nociceptive neurons generate action potentials only when they are stimulated with a noxious stimulus [
5]. Following nerve injury, nociceptive neurons can begin to discharge spontaneously in an abnormal way, and they reduce their response threshold and increase their discharge frequency, evoked by stimulation. These processes are responsible for inducing hyperexcitability of primary sensory neurons (peripheral sensitization) [
5]. In addition, ongoing abnormal activity originating in injured peripheral nerves can be the trigger of central sensitization [
19]. Peripheral and central mechanisms, occurring sequentially and concurrently, are responsible for the induction and maintenance of neuropathic pain [
5].
VGSCs play a role in persistent and chronic pain as it has been reported that changes on VGSC expression in primary sensory neurons, their abnormal accumulation at the site of injury, as well as posttranslational modifications of these channels, occur after nerve injury, and lead to peripheral sensitization processes [
20] (see
Table 1).
Table 1. Implication of TTX-sensitive voltage-gated sodium channels in neuropathic pain.
| Channel |
Normal Localization [16] |
Changes of Expression in Pain States |
| NaV1.1 |
-CNS, PNS-Microglia |
-Unclear after PNI in NP [16] |
| NaV1.2 |
-CNS, very low expression in PNS-SC in lamina I/II |
-Unclear after PNI in NP [16] |
| NaV1.3 |
-Negligible in DRG (embryonic isoform)-SC in lamina I/II |
Upregulated in DRG: CCI [21], PDN [22,23],
SNI [24,25], SNL [26,27],
traumatic nerve injury (human) [28]
Upregulate in painful neuromas (human): [29]
Trigeminal ganglion: trigeminal neuropathic pain [30,31,32]
Upregulated in sciatic nerve: PDN [33]
Upregulated in spinal cord: SCI [34] |
| NaV1.4 |
-Skeletal muscle |
|
| NaV1.6 |
-Nodes of Ranvier-SC-PNS-Epidermal free nerve terminals-keratinocytes-Microglia |
Upregulated in DRG: PDN [35],
lumbar 5 ventral root transection [36],
CINP (oxaliplatin) [37]
Upregulated in sciatic nerve: CCI [38]
Upregulated trigeminal ganglion: trigeminal neuropathic pain [30] |
| NaV1.7 |
-PNS in all types of DRG neurons-SC-Epidermal free nerve terminals |
Downregulated in DRG: SNL [39], SNI [24,25],
traumatic nerve injury (human) [28]
Upregulated in DRG: CCI [40], SNI [41], SNL [27],
CINP (paclitaxel) [42],
cancer-related pain (humans) [42],
Herpesvirus quiescent infection [43],
painful neuromas (human) [34]
Upregulated in spinal cord: CINP (paclitaxel) [42]
Upregulated trigeminal ganglion: trigeminal neuropathic pain [44]
Upregulated in sciatic nerve: CCI [45] |
Thus, it has been shown a massive upregulation of the expression of the TTX-sensitive VGSC Na
V1.3 in peripheral sensory neurons following nerve injury, in several models of neuropathic pain in rodents [
21,
23,
24,
26,
33], as well as in patients [
28,
29]. Na
V1.3 subtype might account for neuronal hyperresponsiveness during neuropathic pain as this isoform is not expressed in peripheral sensory neurons in normal condition (only during embryonic development) [
20], but as mentioned before, it dramatically upregulates following nerve injury in animals and patients with neuropathic pain (see
Table 1). However, specific reduction in the expression of Na
V1.3 subtype alone (with intrathecal administration of antisense oligonucleotides or with knockout mice), is not sufficient to reduce behavioral hypersensitivity associated with nerve injury [
46,
47].
Another TTX-sensitive VGSC that has received much attention is Na
V1.7 subtype. This VGSC is predominantly localized in the peripheral nervous system, and many studies have reported a strong correlation between Na
V1.7 function and many pain states in rodents and humans [
20]. However, the role of Na
V1.7 in neuropathic pain remains uncertain, as contradictory results have been reported in rodents and humans, with some studies showing upregulation [
27,
34,
40,
41,
45], and others downregulation [
24,
28,
39] in primary sensory neurons. In addition, mice lacking Na
V1.7 normally developed neuropathic pain after peripheral nerve injury [
48]. Interestingly, it has been shown that Na
V1.7 could have a more relevant role in the somatosensory system of humans than that of mice, since Na
V1.7 is the predominant VGSC in human dorsal root ganglia (DRG) (50% of total VGSC expression), whereas in mice, this subtype represents only a small percentage (18% of total VGSC expression) [
49]. In addition, in the same study, it is shown that human DRG neurons in primary cultures treated with paclitaxel (an antitumor drug which is known to induce neuropathic pain), increase the expression of Na
V1.7 [
49]. Finally, the importance of Na
V1.7 for pain in humans is revealed by the existence of loss-of-function mutations of Na
V1.7 gene (SCN9A) that lead to congenital insensitivity to pain, and gain-of-function mutations of this gene, which lead to congenital pain disorders characterized by hyperexcitability and extreme pain [
50].
More recently, it has been shown that Na
V1.6 also plays a relevant role in neuropathic pain of different etiologies, as several studies have reported that Na
V1.6 is upregulated during neuropathic pain of different etiologies [
35,
36,
37,
38]. In addition, specific inhibition of Na
V1.6 with small interfering RNA injected into the DRG [
51], or knockout mice [
52], reduce neuropathic pain in rodents. On the other hand, the involvement in neuropathic pain of other TTX-sensitive VGSCs present in the nervous system (Na
V1.1 and Na
V1.2) is not totally clear [
16].
TTX-resistant VGSCs which are preferentially expressed in primary afferent neurons (Na
V1.8 and Na
V1.9) are involved in some pain conditions [
20]. However, the role of these VGSC subtypes in neuropathic pain is also unclear since it has been described that they are downregulated in DRG after nerve injury [
23,
24,
53,
54]. In addition, experiments with knockout mice show a role of Na
V1.9, and to a lesser extent or Na
V1.8 in the generation of cold hypersensitivity, but not mechanical hypersensitivity in models of neuropathic pain [
55]. On the other hand, the principal cardiac VGSC Na
V1.5 is not involved in pain [
17], but it is of particular interest, as any VGSC blocker with analgesic potential, should not inhibit Na
V1.5 at therapeutic doses, for obvious safety reasons. Interestingly, Na
V1.5 is considered a TTX-resistant VGSC [
56].
3. Effects of TTX in Preclinical Models of Neuropathic and Cancer Pain
3.1. Preclinical Studies on Neuropathic Pain
TTX has been widely employed in vitro to characterize the role of VGSCs in nociception and chronic pain. In addition, the effects of TTX on pain behaviors have been studied in several models of acute and chronic pain. Thus, the systemic administration of TTX has shown analgesic effect in classical pain models such as in the writhing test in mice [
57] and rats [
58], and in the formalin test in rats [
57]. In another study, systemic TTX showed a significant reduction of mechanical hyperalgesia in a rat model of inflammatory pain induced by intraplantar carrageenan [
59]. Our group demonstrated that the systemic administration of TTX reduced pain behaviors in different models of visceral pain in mice [
60].
As we have described in the previous section, some TTX-sensitive VGSCs play a key role in neuropathic pain, and this probably explains why the effect of TTX has been more widely tested in preclinical models of neuropathic pain (see Table 2).
Table 2. Summary of the effects of TTX on pain studies in laboratory animals.
| Administration of TTX |
TTX Doses |
Effect (None, Moderate, Strong) |
Pain Test |
Pain Model |
Reference |
| Sciatic nerve blockade |
TTX osmotic pump |
Strong |
MA, TH |
SNI and CCI |
[61] |
| Topical DRG |
12.5–50 nM/5 µL |
Strong (12.5–50 µg) |
MA |
SNL |
[62] |
| Epidural |
25 nM/5 µL |
Strong (25 µg) |
MA |
SNL |
[62] |
| Topical median nerve |
Gel pads with TTX |
Strong |
MA |
CCI |
[63] |
| Intraperitoneal |
25 nM/5 µL |
None |
MA |
SNL |
[62] |
| 8 µg |
None |
MA |
CINP (vincristine) |
[64] |
| Subcutaneous |
1–6 µg |
Strong |
MA, TH, CA |
CINP (paclitaxel) |
[65] |
| 6 µg |
Strong |
MA |
intraplantar capsaicin |
[66] |
| 0.3–6 µg |
Strong (1–6 µg) |
MA, TH |
SNL |
[57] |
| Acute and subchronic TTX (1–6 µg) |
Strong |
MA, TH |
CCI |
[67] |
| Acute and subchronic TTX (1–6 µg) |
Moderate |
MA, TH |
CCI-intraorbital nerve |
[67] |
| 8 µg |
Strong |
MA, TH |
burn-induced pain |
[68] |
| Intragastrical |
5–20 µg |
Strong |
MA, TH |
Postherpetic Neuralgia (RTX) |
[69] |
| Intramuscular |
Acute and subchronic TTX (1–6 µg s.c.) |
Strong |
MA |
Postherpetic Neuralgia (RTX) |
[69] |
| 0.03–1 ug |
Moderate |
MH |
CINP (oxaliplatin) |
[70] |
| Intrathecal |
10 µg |
Strong |
MA, TH |
bone cancer pain |
[71] |
Lyu and collaborators [
62] reported the first study on TTX effects in a model of neuropathic pain in 2000. These authors showed that mechanical allodynia induced by spinal nerve ligation (SNL) was significantly attenuated by the topical application to the DRG of TTX at low doses that did not block action potential conduction. This study also reported that TTX ameliorates mechanical hypersensitivity when applied to the epidural space but not when this neurotoxin was administered intraperitoneally, suggesting a local effect of TTX. However, the dose of TTX used in this study (25 nM) was probably too low to achieve a systemic analgesic effect. In a later study was reported that the nerve blockade with TTX immediately following nerve injury, prevented the development of thermal and mechanical hypersensitivity, and the spontaneous afferent activity, in two rat models of neuropathic pain, the chronic constrictive injury (CCI) and the spared nerve injury (SNI) models [
61]. However, when nerve blockade was performed once the neuropathic pain was established (10 days after nerve injury), it only transiently inhibited mechanical hypersensitivity in both neuropathic pain models, suggesting that the local inhibition of TTX-sensitive VGSCs during the initial stage of neuropathic pain might be sufficient to inhibit the subsequent development of neuropathic pain [
61]. Another report showed that the preventive local application of TTX onto the median nerve inhibited the development of tactile allodynia subsequent to the chronic constriction injury of this nerve, in parallel with a reduction of the increase of astrocyte activation in the cuneate nucleus [
63].
Two independent studies evaluated the effects of systemic administration of TTX in neuropathic pain models induced by surgical nerve damage. Marcil and collaborators [
57] tested the ameliorative effects of several doses of subcutaneous TTX on pain-like behaviors induced by the partial sciatic nerve ligation (pSNL) model in rats and compared such effects with those produced by morphine. They found that TTX reduced mechanical allodynia and thermal hyperalgesia, without affecting the contralateral non-injured side. In contrast, the administration of morphine reduced neuropathic pain, but it also led to an increase in pain threshold strikingly above preoperative values in both ipsilateral and contralateral paws, although the dose of morphine was probably too high. In another report, it was shown that acute and subchronic subcutaneous treatment with TTX suppressed mechanical and thermal hyperalgesia and mechanical allodynia in the rat CCI model [
67]. In the same study, the authors evaluated the effects of subcutaneous TTX on pain behaviors induced by the CCI of the infraorbital nerve in rats, and found that TTX had only modest analgesic effects in this pain condition. They also provided data about the possible mechanisms that contribute to the ameliorative effects of TTX in neuropathic pain. In particular, they showed that acute subcutaneous administration of TTX attenuated the increase of c-Fos expression (widely used as marker of neuronal activity) in the dorsal horn of the spinal cord and in some supraspinal areas in response to light mechanical stimulation of the ipsilateral hind paw. In addition, they found that TTX alleviated pain-related behaviors induced by nerve injury through mechanisms that might involve complex interactions with opioidergic systems, whereas noradrenergic and serotoninergic systems were not involved [
67].
TTX was also evaluated in several models of cancer chemotherapy-induced neuropathic pain with contradictory results. Thus, the intraperitoneal administration of TTX did not attenuate mechanical allodynia induced by vincristine in rats, whereas other drugs including morphine, pregabalin, mexiletine and lidocaine did [
64]. In a more recent study, Alvarez and Levine [
70] showed that intramuscular TTX produced a small but significant reduction of neuropathic muscle pain induced by the antineoplastic agent oxaliplatin in rats. Our group found that acute TTX administered subcutaneously attenuated mechanical and cold allodynia and heat hyperalgesia induced by paclitaxel in mice, once the neuropathy was already established [
65]. In addition, we also showed that the preventative administration of TTX completely prevented the development of mechanical and cold allodynia, without affecting heat hyperalgesia. Such findings open the possibility of preventative treatment of neuropathic pain induced by chemotherapy in patients, where the moment of onset of the nerve insult is known and preventive treatment could be possible. The mechanism by which TTX prevents chemotherapy-induced neuropathic pain is unknown. However, it has been shown that chemotherapy-induced neuropathy is associated with upregulation of some TTX-sensitive VGSCs in the DRG and in the spinal cord (see
Table 2). It is possible to speculate that blockade of the early spontaneous afferent activity generated by the upregulation of TTX-sensitive VGSCs during the development of chemotherapy-induced neuropathic pain explains the preventive effect of TTX. A similar mechanism has been shown in other models of neuropathic pain [
61]. On the other hand, the discrepancies between the results obtained by TTX in the models of neuropathic pain induced by three different chemotherapeutic agents may be attributable to the different underlying mechanisms by which these anticancer drugs induce neuropathic pain [
72].
Other studies evaluated the effects of systemic administration of TTX in preclinical models of neuropathic pain of different etiologies other than those induced by a surgical lesion or chemotherapy. Thus, the subcutaneous administration of TTX suppressed thermal hyperalgesia and mechanical allodynia in a rat full burn wound pain model. Interestingly, the results of this study indicated that TTX reduced thermal hyperalgesia more effectively than morphine, whereas both drugs showed similar effects reducing mechanical allodynia [
68]. More recently, the effects of oral administration of TTX pellets have been evaluated in a rat model of postherpetic neuralgia [
69]. In this study, intragastric treatment with TTX was able to reduce mechanical allodynia in a similar way than oral pregabalin. In addition, these authors also showed that the intramuscular administration with TTX reduced mechanical allodynia at doses significantly lower than those used during the oral treatments. However, they claimed the TTX pellets described in this article might have a higher therapeutic window than the intramuscular administration of TTX (used in patients). It would be interesting to evaluate this new pharmaceutical form of TTX in more widely used models of neuropathic pain than the unusual model of postherpetic neuralgia induced by resiniferatoxin (RTX). Finally, our group reported that TTX inhibited capsaicin-induced mechanical hypersensitivity in mice [
66], which is considered a surrogate model of neuropathic pain [
73].
In summary, the systemic or local treatment with TTX has shown efficacy in reducing pain-like behaviors in several animal models of neuropathic pain of different etiology. There is only one study that, by using an adequate systemic dose of TTX, failed to inhibit neuropathic pain in rodents [
64]. Therefore, these findings point to the therapeutic usefulness of TTX in neuropathic pain and support the key role that TTX-sensitive VGSCs play in neuropathic pain states (see previous section). Nevertheless, there is still much to learn about the effects of this drug at the preclinical level. In fact, the analgesic effect of TTX in certain preclinical models that mimic highly prevalent painful neuropathies, such as diabetic neuropathy, has never been explored.
3.2. Preclinical Studies on Cancer Pain
Chronic cancer-related pain encompasses mixed pathophysiology as a consequence of tissue response to the primary tumor or metastases, and it can be considered a type of mixed nociceptive and neuropathic pain [
74]. Although TTX has been used in several clinical trials to relieve cancer-related pain in patients (see next section), it is surprising that this drug is almost unexplored in preclinical models of pain induced by cancer, with only one article. Zhen and collaborators [
71] evaluated the role of TTX in bone cancer pain. They found that sustained intrathecal injection of TTX (10 μg/kg, once a day) significantly attenuated mechanical allodynia and thermal hyperalgesia in rats with bone cancer.
With only one preclinical study, it seems clear that more research is needed to evaluate the effects of TTX in models of cancer-related pain.