The quality of honey bee drone semen is relevant in different contexts, ranging from colony productivity to pathology, toxicology and biodiversity preservation. Despite its importance, considerably less knowledge is available on this subject for the honey bee when compared to other domestic animal species. A proper assessment of sperm quality requires a multiple testing approach which discriminates between the different aspects of sperm integrity and functionality. Most studies on drone semen quality have only assessed a few parameters, such as sperm volume, sperm concentration and/or sperm plasma membrane integrity.
- Apis mellifera
- male
- reproduction
- semen
- sperm quality
1. Introduction
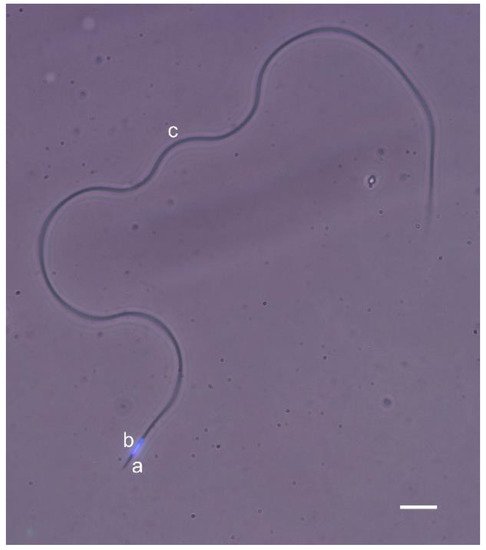
2. Normal Sperm Structure in the Honey Bee
3. Sperm Life Cycle in the Honey Bee
3.1. Spermatogenesis and Sperm Storage in the Male
3.2. Mating, Sperm Storage in the Spermatheca and Egg Fertilization
This entry is adapted from the peer-reviewed paper 10.3390/biology9070174
References
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353.
- Soroker, V.; Hetzroni, A.; Yakobson, B.; David, D.; David, A.; Voet, H.; Slabezki, Y.; Efrat, H.; Levski, S.; Kamer, Y.; et al. Evaluation of colony losses in Israel in relation to the incidence of pathogens and pests. Apidologie 2011, 42, 192–199.
- Clermont, A.; Eickermann, M.; Kraus, F.; Georges, C.; Hoffmann, L.; Beyer, M. A survey on some factors potentially affecting losses of managed honey bee colonies in Luxembourg over the winters 2010/2011 and 2011/2012. J. Apic. Res. 2014, 53, 43–56.
- Dainat, B.; Evans, J.D.; Chen, Y.P.; Gauthier, L.; Neumann, P. Predictive markers of honey bee colony collapse. PLoS ONE 2012, 7, e32151.
- Pettis, J.S.; Rice, N.; Joselow, K.; van Engelsdorp, D.; Chaimanee, V. Colony failure linked to low sperm viability in honey bee (Apis mellifera) queens and an exploration of potential causative factors. PLoS ONE 2016, 11, e0147220.
- Collins, A.M. Relationship between semen quality and performance of instrumentally inseminated honey bee queens. Apidologie 2000, 31, 421–429.
- Collins, A.M. Functional longevity of honey bee, Apis mellifera, queens inseminated with low viability semen. J. Apic. Res. 2004, 43, 167–171.
- Tarpy, D.R.; Olivarez, R. Measuring sperm viability over time in honey bee queens to determine patterns in stored-sperm and queen longevity. J. Apic. Res. 2014, 53, 493–495.
- Locke, S.J.; Peng, Y.S. The effects of drone age, semen storage and contamination on semen quality in the honey-bee (Apis mellifera). Physiol. Entomol. 1993, 18, 144–148.
- Rhodes, J.W.; Harden, S.; Spooner-Hart, R.; Anderson, D.L.; Wheen, G. Effects of age, season and genetics on semen and sperm production in Apis mellifera drones. Apidologie 2011, 42, 29–38.
- Stürup, M.; Baer-Imhoof, B.; Nash, D.R.; Boomsma, J.J.; Baer, B. When every sperm counts: Factors affecting male fertility in the honeybee Apis mellifera. Behav. Ecol. 2013, 24, 1192–1198.
- Rousseau, A.; Fournier, V.; Giovenazzo, P. Apis mellifera (Hymenoptera: Apidae) drone sperm quality in relation to age, genetic line, and time of breeding. Can. Entomol. 2015, 147, 702–711.
- Schluns, H.; Schluns, E.A.; van Praagh, J.; Moritz, R.F.A. Sperm numbers in drone honeybees (Apis mellifera) depend on body size. Apidologie 2003, 34, 577–584.
- Czekonska, K.; Chuda-Mickiewicz, B.; Chorbinski, P. The effect of brood incubation temperature on the reproductive value of honey bee (Apis mellifera) drones. J. Apic. Res. 2013, 52, 96–105.
- Abdelkader, F.B.; Kairo, G.; Tchamitchian, S.; Cousin, M.; Senechal, J.; Crauser, D.; Vermandere, J.P.; Alaux, C.; Conte, Y.L.; Belzunces, L.P.; et al. Semen quality of honey bee drones maintained from emergence to sexual maturity under laboratory, semi-field and field conditions. Apidologie 2014, 45, 215–223.
- Czekonska, K.; Chuda-Mickiewicz, B.; Samborski, J. Quality of honeybee drones reared in colonies with limited and unlimited access to pollen. Apidologie 2015, 46, 1–9.
- Zaitoun, S.; Al-Ghzawi, A.A.M.; Kridli, R. Monthly changes in various drone characteristics of Apis mellifera ligustica and Apis mellifera syriaca. Entomol. Sci. 2009, 12, 208–214.
- Collins, A.M.; Pettis, J.S. Effect of varroa infestation on semen quality. Am. Bee J. 2001, 141, 590–593.
- DelCacho, E.; Marti, J.I.; Josa, A.; Quilez, J.; Sanchez-Acedo, C. Effect of Varroa jacobsoni parasitization in the glycoprotein expression on Apis mellifera spermatozoa. Apidologie 1996, 27, 87–92.
- Ciereszko, A.; Wilde, J.; Dietrich, G.J.; Siuda, M.; Bak, B.; Judycka, S.; Karol, H. Sperm parameters of honeybee drones exposed to imidacloprid. Apidologie 2017, 48, 211–222.
- Gajger, I.T.; Sakac, M.; Gregorc, A. Impact of thiamethoxam on honey bee queen (Apis mellifera carnica) reproductive morphology and physiology. Bull. Environ. Contam. Toxicol. 2017, 99, 297–302.
- Johnson, R.M.; Dahlgren, L.; Siegfried, B.D.; Ellis, M.D. Effect of in-hive miticides on drone honey bee survival and sperm viability. J. Apic. Res. 2013, 52.
- Taylor, M.A.; Guzman-Novoa, E.; Morfin, N.; Buhr, M.M. Improving viability of cryopreserved honey bee (Apis mellifera L.) sperm with selected diluents, cryoprotectants, and semen dilution ratios. Theriogenology 2009, 72, 149–159.
- Hopkins, B.K.; Herr, C. Factors affecting the successful cryopreservation of honey bee (Apis mellifera) spermatozoa. Apidologie 2010, 41, 548–556.
- Wegener, J.; May, T.; Knollmann, U.; Kamp, G.; Muller, K.; Bienefeld, K. In vivo validation of in vitro quality tests for cryopreserved honey bee semen. Cryobiology 2012, 65, 126–131.
- Hopkins, B.K.; Cobey, S.W.; Herr, C.; Sheppard, W.S. Gel-coated tubes extend above-freezing storage of honey bee (Apis mellifera) semen to 439 days with production of fertilised offspring. Reprod. Fertil. Dev. 2017, 29, 1944–1949.
- Collins, A.M. A scientific note on the effect of centrifugation on pooled honey bee semen. Apidologie 2003, 34, 469–470.
- Collins, A.M. Sources of variation in the viability of honey bee, Apis mellifera L., semen collected for artificial insemination. Invertebr. Reprod. Dev. 2004, 45, 231–237.
- Shafir, S.; Kabanoff, L.; Duncan, M.; Oldroyd, B.P. Honey bee (Apis mellifera) sperm competition in vitro—Two are no less viable than one. Apidologie 2009, 40, 556–561.
- Den Boer, S.P.; Baer, B.; Boomsma, J.J. Seminal fluid mediates ejaculate competition in social insects. Science 2010, 327, 1506–1509.
- Lino-Neto, J.; Bao, S.N.; Dolder, H. Sperm ultrastructure of the honey bee (Apis mellifera) (L) (Hymenoptera, Apidae) with emphasis on the nucleus-flagellum transition region. Tissue Cell 2000, 32, 322–327.
- Phillips, D.M. Insect sperm: Their structure and morphogenesis. J. Cell Biol. 1970, 44, 243.
- Peng, C.Y.S.; Yin, C.M.; Yin, L.R.S. Ultrastructure of honey-Bee, Apis mellifera, sperm with special emphasis on the acrosomal complex following high-pressure freezing fixation. Physiol. Entomol. 1993, 18, 93–101.
- Bishop, G.H. Fertilization in the honey-bee. I. The male sexual organs: Their histological structure and physiological functioning. J. Exp. Zool 1920, 31, 224–265.
- Hoage, T.R.; Kessel, R.G. An electron microscope study of the process of differentiation during spermatogenesis in the drone honey bee (Apis mellifera L.) with special reference to centriole replication and elimination. J. Ultrastruct. Res. 1968, 24, 6–32.
- Jaycox, E.R. The Effects of Various Foods and Temperatures on Sexual Maturity of the Drone Honey Bee (Apis mellifera). Ann. Entomol. Soc. Am. 1961, 54, 519–523.
- Hayashi, S.; Satoh, T. Sperm maturation process occurs in the seminal vesicle following sperm transition from testis in honey bee males. Apidologie 2019, 50, 369–378.
- Couvillon, M.J.; Hughes, W.O.H.; Perez-Sato, J.A.; Martin, S.J.; Roy, G.G.F.; Ratnieks, F.L.W. Sexual selection in honey bees: Colony variation and the importance of size in male mating success. Behav. Ecol. 2010, 21, 520–525.
- Hellmich, R.L.; Rinderer, T.E.; Danka, R.G.; Collins, A.M.; Boykin, D.L. Flight Times of Africanized and European Honey-Bee Drones (Hymenoptera, Apidae). J. Econ. Entomol. 1991, 84, 61–64.
- Reyes, M.; Crauser, D.; Prado, A.; Le Conte, Y. Flight activity of honey bee (Apis mellifera) drones. Apidologie 2019, 50, 669–680.
- Baudry, E.; Solignac, M.; Garnery, L.; Gries, M.; Cornuet, J.M.; Koeniger, N. Relatedness among honeybees (Apis mellifera) of a drone congregation. Proc. Biol. Sci. 1998, 265, 2009–2014.
- Schluns, H.; Moritz, R.F.A.; Neumann, P.; Kryger, P.; Koeniger, G. Multiple nuptial flights, sperm transfer and the evolution of extreme polyandry in honeybee queens. Anim. Behav. 2005, 70, 125–131.
- Koeniger, N.; Koeniger, G. Mating flight duration of Apis mellifera queens: As short as possible, as long as necessary. Apidologie 2007, 38, 606–611.
- Taber, S.; Wendel, J. Concerning the number of times queen bees mate. J. Econ. Entomol. 1958, 51, 786–789.
- Kraus, F.B.; Koeniger, N.; Tingek, S.; Moritz, R.F.A. Using drones for estimating colony number by microsatellite DNA analyses of haploid males in Apis. Apidologie 2005, 36, 223–229.
- Withrow, J.M.; Tarpy, D.R. Cryptic “royal” subfamilies in honey bee (Apis mellifera) colonies. PLoS ONE 2018, 13, e0199124.
- Brutscher, L.M.; Baer, B.; Nino, E.L. Putative Drone Copulation Factors Regulating Honey Bee (Apis mellifera) Queen Reproduction and Health: A Review. Insects 2019, 10, 8.
- Haberl, M.; Tautz, D. Sperm usage in honey bees. Behav. Ecol. Sociobiol. 1998, 42, 247–255.
- Baer, B. Sexual selection in Apis bees. Apidologie 2005, 36, 187–200.
- Laidlaw, H.H.; Page, R.E. Polyandry in honey bees (Apis-mellifera L)—Sperm utilization and intracolony genetic-relationships. Genetics 1984, 108, 985–997.
- Tarpy, D.R.; Keller, J.J.; Caren, J.R.; Delaney, D.A. Assessing the mating ‘health’ of commercial honey bee queens. J. Econ. Entomol. 2012, 105, 20–25.
- Lodesani, M.; Balduzzi, D.; Galli, A. A study on spermatozoa viability over time in honey bee (Apis mellifera ligustica) queen spermathecae. J. Apic. Res. 2004, 43, 27–28.
- Baer, B.; Collins, J.; Maalaps, K.; den Boer, S.P. Sperm use economy of honeybee (Apis mellifera) queens. Ecol. Evol. 2016, 6, 2877–2885.