(1) In a cafeteria experiment, beavers chose P. fremontii six times more often than other woody native and exotic species. (2) With two cottonwood species, we found that the nitrogen and salicortin concentrations were up to 45% greater and lignin concentration 14% lower in the juvenile resprout growth of felled trees than the juvenile growth on unfelled trees (six of seven analyses were significant for P. fremontii and four of six were significant for P. angustifolia). (3) With two cottonwood species, arthropod community composition on juvenile branches differed significantly between felled and unfelled trees, with up to 38% greater species richness, 114% greater relative abundance and 1282% greater species diversity on felled trees (six of seven analyses with P. fremontii and four of six analyses with P. angustifolia were significant). The above findings indicate that the highest arthropod diversity is achieved in the heterogenous stands of mixed felled and unfelled trees than in stands of cottonwoods, where beavers are not present. These results also indicate that beaver herbivory changes the chemical composition in 10 out of 13 chemical traits in the juvenile growth of two of the three cottonwood species to potentially allow better defense against future beaver herbivory. (4) With P. deltoides, only one of five analyses in chemistry was significant, and none of the four arthropod community analyses were significant, suggesting that this species and its arthropod community responds differently to beaver. Potential reasons for these differences are unknown. Overall, our findings suggest that in addition to their impact on riparian vegetation, other mammals, birds, and aquatic organisms, beavers also may define the arthropod communities of two of three foundation tree species in these riparian ecosystems.
1. Introduction
The ecological processes that structure communities are fundamental aspects of ecology and evolution. Holling [
1] proposed that “a small set of plant, animal, and abiotic processes structure ecosystems across scales in time and space”. In other words, not all species are equal, and a few are likely to contribute disproportionately to the structure and evolution of communities and ecosystems. Species that are strong interactors, affecting many other species and modifying their environments (e.g., dam building and selective foraging by beavers; shading and cooling of the understory by tree canopies) likely play a major role in structuring communities. Dayton [
2] defined a foundation species as “a single species that defines much of the structure of a community by creating locally stable conditions for other species, and by modulating and stabilizing fundamental ecosystem processes”. Ellison et al. [
3] reviewed this literature and concluded that dominant species, keystone species, ecosystem engineers and other strong interactors all fall within this category of foundation species. Because all ecosystems of the world likely have multiple and potentially interacting foundation species, it is especially important to identify and understand the interactions of these species as their loss due to global change (including climate change, invasive species, and altered species interactions) could cascade to affect the rest of the community [
4]. Keith et al. [
5,
6] proposed and experimentally tested the interacting foundation species hypothesis and found that the interactions of two foundation species better explained community diversity, stability and species interaction networks than either one alone.
Beavers and cottonwoods are considered foundation species because their presence in an ecosystem has far-reaching effects on community dynamics and ecosystem processes [
3]. By felling trees that are used for food and construction activities (e.g., building dams that slow water flow and create ponds and lodges), beavers affect fish populations [
7], stream flow and water temperature [
8,
9], nutrient cycling and availability [
9,
10,
11], individual species and communities of arboreal arthropods [
12,
13,
14], the composition of mixed cottonwood riparian forests and their genetic composition [
15], communities of aquatic arthropods [
16], vegetation diversity [
17], tree architecture [
18], and genetic diversity and productivity at the individual tree level [
19]. Even in areas where beavers are no longer active but have cut trees and built dams in the recent past, their impact is evident in the structure of the riparian zone and has long-term effects on vegetation types [
9,
11,
20,
21,
22,
23]. Compared with the amount of land they cover, riparian corridors contain disproportionately high biodiversity [
24] and are an important resource for birds and mammals. In the arid southwest USA, the narrow riparian “ribbon of green” (
Figure 1A) has been shown to support exceptionally high bird [
25] and vegetation diversity [
26]. However, the extent to which beavers interact with a foundation tree species to affect arthropod diversity in these southwestern stream systems has not been studied (but see [
14]).
Figure 1. (A) Riparian “ribbon of green” of P. fremontii near Moab, UT; (B) Resprout growth of P. angustifolia after felling by beavers; (C) Leaf rolling moth (Anacampsis niveopulvella Chambers, 1875; an indicator species of resprout growth after P. fremontii felling by beavers); (D) Leaf rolling moth larvae, a member of the arthropod community influenced by beaver herbivory, in the family Tortricidae (also indicator species of resprout growth) create habitat for other arthropods, such as this spider.
One mechanism by which beaver herbivory might affect arthropod diversity is by altering the chemistry of the plants they browse. To determine whether there was a difference in twig chemistry between the juvenile resprout growth of beaver-felled trees (
Figure 1B), and the juvenile shoots at the base of unfelled trees, we collected samples from each to assess the concentrations of a suite of chemicals known to be present in these tissues. Since beavers eat the inner bark, or cambium layer of trees [
27], we chose to sample twig tissue to quantify how beaver herbivory might affect the phytochemistry of resprouting cottonwoods for future feeding by beavers. While we cannot extrapolate on how twig chemistry may influence foliar arthropods, we were also interested in understanding how herbivory by beavers may differentially affect arthropods on multiple cottonwood species, which has not previously been examined in this system.
To examine how beaver herbivory on Fremont cottonwood (P. fremontii), Eastern cottonwood (P. deltoides) and narrowleaf cottonwood (P. angustifolia) in different southwestern river systems may change twig chemistry and arthropod communities (see representative arthropod species Figure 1C,D), we tested three hypotheses: (1) Beavers show a preference for Fremont cottonwood over three other native species and one invasive woody species common to the area. (2) The resprouted juvenile stems of unfelled trees and trees felled by beavers differ in their levels of nitrogen, carbon, lignin, condensed tannins and the phenolic glycosides salicortin and HCH-salicortin. (3) The arthropod communities in paired samples of juvenile foliage on felled and unfelled trees differ in their multivariate community parameters including metrics such as species richness and abundance.
Testing these hypotheses will increase our knowledge of how the interactions of plant and mammalian foundation species impact a diverse arthropod community. While several studies have examined the impacts of beavers on aquatic arthropods [
16] and single arboreal arthropod species [
12,
13], only one other study that we are aware of examined the impacts of beaver felling of trees on a diverse arboreal arthropod community on narrowleaf cottonwood in another region [
14]. Understanding the interactions among ecosystem-defining species (beavers, different species of cottonwoods and other native and exotic trees/shrubs), phytochemistry, and arthropod communities will allow for more holistic approaches to management and conservation in these fragile southwest riparian habitats.
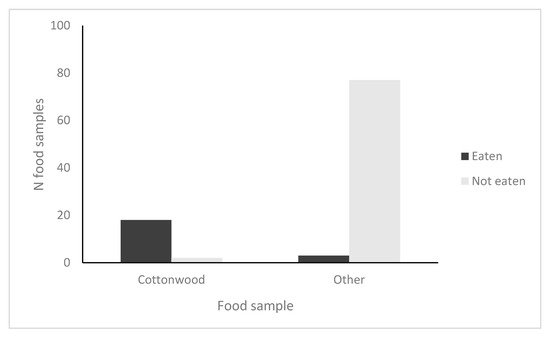
2. Beaver Cafeteria Feeding Experiment
In trial 1 (collected after three days), 80% of cottonwood branches were eaten but no branches were eaten of tamarisk, sycamore, ash or willow. In trial 2 (collected after 10 days), 100% of cottonwood branches were eaten, 10% (1/10) and 20% (2/10) of the branches of ash and willow were eaten, respectively, and no branches of tamarisk and sycamore were eaten. To meet R × C
G-test assumptions [
47], we collapsed the results of the trials into a matrix with two rows (eaten; not eaten) and two columns (cottonwood; non-cottonwood). The resulting 2 × 2
G-test showed a significant preference by beavers for cottonwood branches over the other available woody vegetation (
G = 64.2, df = 1,
p < 0.0001,
N = 100;
Figure 2).
Figure 2. Results of foraging preferences for beavers at Dry Beaver Creek, AZ in four trials of 25 (100 total samples). Beavers ate 18/20 (90%) of cottonwood samples and 3/80 (4%) of non-cottonwood samples; the data collapsed into a 2 × 2 table for analysis were significant (G = 64.2, df = 1, p < 0.0001).
3. Tree Phytochemical Responses to Beaver Herbivory
Arizona Sites: Our MRPP/NMDS analyses of Fremont cottonwood trees indicated that total twig chemistry differed between study locations as well as between the juvenile resprout growth of felled and the juvenile growth of unfelled Fremont cottonwoods. For the Arizona study, with 24 pairs of trees, we found significant effects for Site (Dry Beaver Creek, Verde River; A = 0.125; p < 0.0001) and Treatment (Felled, Unfelled; A = 0.016, p = 0.0163; Figure 3A) and a significant Site × Treatment interaction (A = 0.144, p < 0.001). Separate MRPP/NMDS analysis of twig chemistry for each AZ site (Bonferroni-adjusted α = 0.05/2 = 0.025) showed a significant chemical difference between felled and unfelled trees at Dry Beaver Creek (DBC; A = 0.025, p = 0.019), but no significant chemical differences between similar trees at Verde River (VR; A = 0.017, p = 0.063). Samples from resprout growth on felled Fremont cottonwood at Arizona sites showed 14% higher nitrogen concentration than those of the juvenile growth of unfelled trees (p < 0.0001). Our Bonferroni-adjusted pairwise Wilcoxon signed-rank tests for five phytochemicals carbon, nitrogen, lignin, salicortin and HCH-salicortin; adjusted α = 0.05/(5 + 2) = 0.007) showed significant differences between felled and unfelled trees for nitrogen (p = 0.0001), lignin (p = 0.0035), salicortin (p = 0.002) and HCH-salicortin (p = 0.009) at Dry Beaver Creek only. We found no significant differences between felled and unfelled trees for carbon (p = 0.10) or for HCH-salicortin at the Verde River (p = 0.035).
Figure 3. Non-metric multidimensional scaling (NMDS) compared chemistry profiles of juvenile felled and unfelled cottonwood twigs: (A) Fremont cottonwoods in Arizona had significant effects for Site (Dry Beaver Creek, Verde River; A = 0.125; p < 0.0001) and Treatment (Figure shown: Felled, Unfelled; A = 0.016, p = 0.0163) and a significant Site × Treatment interaction (A = 0.144, p < 0.001). (B) In narrowleaf cottonwoods on the San Miguel River, there was also a significant difference between the chemical composition of resprout growth and twigs from unfelled trees (A = 0.123, p < 0.0001); (C) Eastern cottonwoods on the San Miguel River, CO showed no significant difference in chemistry profiles between felled and unfelled trees (A = −0.005, p = 0.59). Icons represent felled and unfelled profiles; centroids are mean values with 95% confidence intervals.
4. Arthropod Responses to Herbivory
Arizona Sites: Our MRPP/NMDS analyses of Fremont cottonwood trees indicated that arthropod communities differ between study locations as well as between the juvenile resprout growth of felled and unfelled Fremont cottonwoods. We found significant differences in the arthropod communities on one collection date (August 2009). We also found significant Site, Treatment and Site x Treatment effects in both sampling dates (Site: AZ May 2009: A = 0.017; p < 0.0001; August 2009: A = 0.023; p < 0.0001; Treatment: Felled, Unfelled: AZ May 2009: A = 0.013; p < 0.005; August 2009: A = 0.012; p < 0.0001; Site × Treatment: AZ May 2009: A = 0.022; p < 0.0001; August 2009: A = 0.036; p < 0.0001). We considered separate MRPP/NMDS analysis of arthropod communities on felled and unfelled trees for each AZ site unjustified, given the unusual rigor of our Bonferroni-adjusted criterion for this comparison (α = 0.05/6 = 0.008).
For Fremont cottonwood in Arizona, our hypothesis that arthropod species richness and abundance would differ significantly between felled and unfelled trees at each site was also supported. For the August 2009 sampling period, arthropod species richness was 56.9% greater in felled trees at Dry Beaver Creek than in unfelled trees. Arthropod richness at the Verde River showed this same pattern in August 2009, with felled trees supporting 38.3% greater arthropod richness than unfelled trees. Relativized arthropod abundance on resprout growth of felled trees was 92.3% greater at Dry Beaver Creek and 114.4% greater on the Verde River than relativized abundance on juvenile growth of unfelled trees in August 2009. Total abundance did not show significant differences. Shannon’s Diversity at Dry Beaver Creek was 1282% higher on felled than on unfelled trees, and at the Verde River, Shannon’s Diversity was 31.7% higher on the resprout growth of felled trees than on paired unfelled trees. With the rigor of Bonferroni adjustments, the May 2009 sampling period only showed a significant difference with respect to Shannon’s Diversity, with felled trees at the Dry Beaver Creek site supporting 52% higher species diversity than unfelled trees. For all other metrics, there was no significant difference between treatments, though 3 analyses (total abundance at the Verde River; relativized abundance and species richness at Dry Beaver Creek) were significant before Bonferroni adjustments. Importantly, there were no cases of the juvenile growth of unfelled trees supporting significantly greater richness, abundance or diversity than resprout growth on felled trees.
Indicator species—We found three arthropod species served as indicator species of beaver-felled trees in Arizona Fremont cottonwoods, and one species that was an indicator of unfelled trees in Colorado narrowleaf cottonwoods. At the Verde River in August 2009, two leaf modifiers were indicative of resprout growth on felled trees: Anacampsis niveopulvella Chambers, 1875 (order Lepidoptera, family Gelichiidae, Figure 1B) had an indicator value of 33.2% (p = 0.0286), and a species of clearwing moth in the order Lepidoptera and family Sesiidae had an indicator value of 41.6% (p = 0.0412). At Dry Beaver Creek, a leafroller moth in the order Lepidoptera and family Tortricidae (Figure 1C) had an indicator value of 27.8% (p = 0.0488) on beaver-felled trees.
Colorado Sites: Our MRPP/NMDS analyses of narrowleaf and Eastern cottonwood trees on the San Miguel River supported the hypothesis that arthropod communities would differ between study locations as well as between the juvenile resprout growth of felled and unfelled trees for narrowleaf cottonwoods (A = 0.015, p = 0.0002), but not for Eastern cottonwoods (A = −0.00099, p = 0.52).
For narrowleaf cottonwood on the San Miguel River, surveys again showed that beaver herbivory generally had a positive impact on arthropod abundance (total abundance, p = 0.004), but no significant difference in species richness (p = 0.038, significant prior to Bonferroni adjustment) or Shannon’s Diversity (p = 0.20). Resprout growth on felled narrowleaf trees supported 115.1% higher total abundance than juvenile growth on unfelled trees. We found no evidence in narrowleaf of unfelled trees supporting significantly greater arthropod richness or abundance than resprout growth on felled trees.
Indicator species—In narrowleaf cottonwood on the San Miguel River, one species of ant (Formica propinqua Creighton, 1940, order Hymenoptera, family Formicidae) was indicative of unfelled trees, with an indicator value of 58.3% (p = 0.0122).
5. Conclusions
Our study found patterns largely in agreement with our hypotheses that beavers preferentially choose cottonwoods over other tree species, impacting cottonwood twig chemistry and influencing arboreal arthropod communities: (1) When offered a variety of locally occurring woody plant species, beavers preferentially selected Fremont cottonwood six times more often than other available food samples; (2) Twig tissue from resprout growth of felled trees showed different defensive, structural and nutritional phytochemistry concentrations than twig tissue from paired unfelled trees; and (3) Juvenile resprout growth on beaver-felled trees supported significantly different arthropod communities, often with higher species richness, diversity and abundance than the juvenile growth of unfelled cottonwoods. This trio of patterns was repeated in both Fremont and narrowleaf cottonwood stands, arguing that cottonwood stands with heterogeneity resulting from beaver felling support higher arthropod diversity than cottonwood stands without beaver influence. These alterations to the phytochemistry of juvenile resprout growth following beaver felling may impact future herbivory. Strikingly, Eastern cottonwood did not follow these patterns, and we are unaware of studies that have investigated reasons for this different response. Our findings indicate that as interacting foundation species, beavers and cottonwoods can impact the phytochemistry of riparian tree stands as well as the community structure of arthropods on these trees. As riparian habitat becomes ever more imperiled due to climate change, invasive species, and development, our understanding of these interactions and their community-wide impacts are increasingly critical for conservation and management. As we further our understanding of these complex and fragile ecosystems, we can be better prepared to prioritize research and responses that protect and restore functionality for biotic communities and ecosystem processes.
This entry is adapted from the peer-reviewed paper 10.3390/f12070877