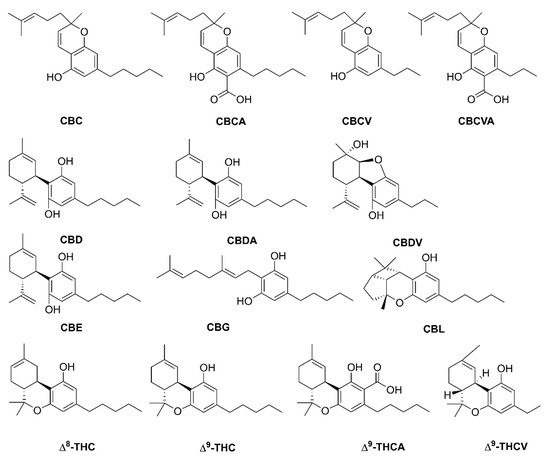
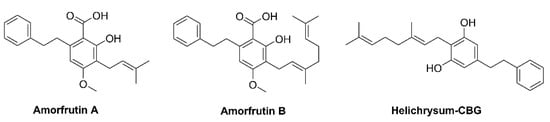
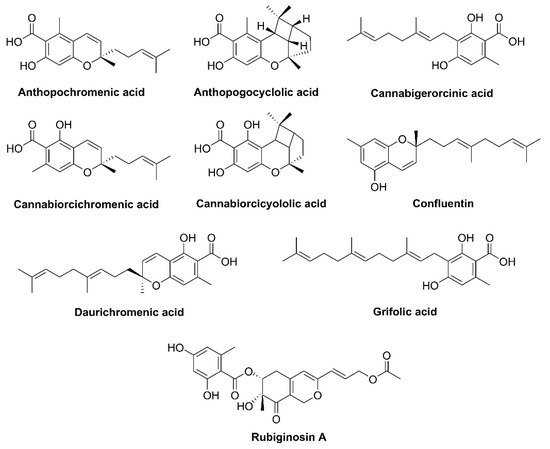
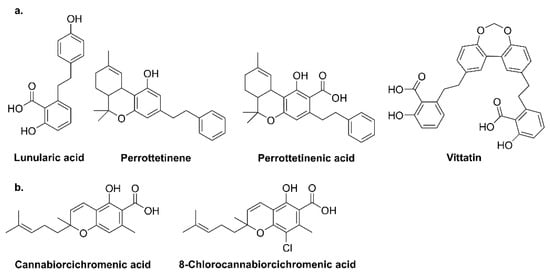
1. Introduction
Phytocannabinoids are meroterpenoids bearing a resorcinol core with an isoprenyl, alkyl, or aralkyl para-positioned side chain, or alkyl group usually containing an odd number of carbon atoms—cannabinoids that have an even number of carbon atoms in the side chain are rare. Phytocannabinoids can be obtained from angiosperms (flowering plants), fungi, and liverworts (
Figure 1). The first phytocannabinoid was isolated from the
Cannabis sativa family Cannabaceae, but it has a long controversial history of its use and abuse [
1,
2]. From
C. sativa more than 113 phytocannabinoids were isolated and classified into several groups such as cannabidiols (CBDs), cannabigerols (CBGs), cannabicyclols (CBLs), cannabidiols (CBNDs), cannabinols (CBNs), cannabitriols (CBTs), cannabichromenes (CBCs), (−)-Δ
9-
trans-tetrahydrocannabinol (Δ
9-THC) and miscellaneous cannabinoids [
1,
3,
4,
5]. Compounds obtained from
C. sativa predominately generate alkyl-type phytocannabinoids with a monoterpene isoprenyl and the pentyl side chain [
4,
6]. In
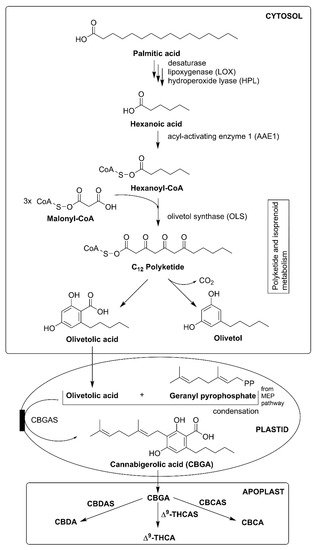
C. sativa, CBD, CBG, CBC, cannabichromevarine (CBCV), and Δ
9-THC are the most abundant cannabinoids in their respective acidic form. The acidic form of the cannabinoid (C22, “pre-cannabinoids”) is the final step of the cannabinoid biosynthetic pathway. Oxidation, decarboxylation, and cyclization lead to the development of modified phytocannabinoid via spontaneous breakdown or conversion product. The conversion mainly occurs due to the poor oxidative stability of phytocannabinoids, especially with the alkyl group.
C. sativa produces the most common phytocannabinoids. In addition to this, the brains of mammals have receptors that respond to the
C. sativa cannabinoid, so they were termed as cannabinoid receptor types 1 and 2 (CB
1R and CB
2R) and thus participated in the endocannabinoid system [
1,
3,
4,
7,
8].
Figure 1. Structure of phytocannabinoids in Cannabis sativa. Abbreviations: CBC, cannabichromene; CBCA, cannabichromenic acid; CBCV, cannabichromevarine; CBCVA, cannabichromevarinic acid; CBD, cannabidiol; CBDA, cannabidiolic acid; CBDV, cannabidivarine, CBE, cannabielsoin; CBG, cannabigerol; CBL, cannabicyclol; Δ8-THC, Δ8-tetrahydrocannabinol; Δ9-THC, Δ9-tetrahydrocannabinol; Δ9-THCA, Δ9-tetrahydrocannabinolic acid; Δ9-THCV, Δ9-tetrahydrocannabivarinic acid.
The Endocannabinoid system in humans and animals revealed that it participates in the regulation of biological functions such as memory, brain system, mood and addiction along with cellular and metabolic processes such as glycolysis, lipolysis, and the energy balance system [
7,
9]. Other angiosperms such as
Helichrysum umbraculigerum (Asteraceae) native to South Africa,
Amorpha fruticosa (Fabaceae), and
Glycyrrhiza foetida (Fabaceae) contains a bioactive compound bearing a cannabinoid backbone (
Figure 2); they are characterized as prenylated bibenzyl derivatives because the aralkyl side chain occurs [
1,
10].
Figure 2. Structure of phytocannabinoids in Helichrysum and Glycyrrhiza plants.
Many
Rhododendron species (family Ericaceae) such as
Rh. dauricum native to Northeastern Asia,
Rh. adamsii found in Eastern Siberia and Mongolia,
Rh. rubiginosum var.
rubiginosum native to Southwest China, and
Rh. anthopogonoides grown in Southern China, all generate active monoterpenoids that have a cannabinoid backbone. Phytocannabinoids are CBC types with an orcinol or methyl group side chain (
Figure 3).
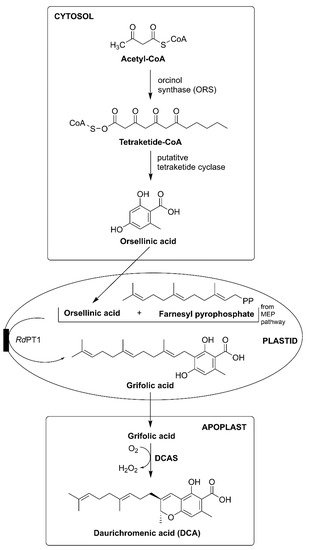
Rh. dauricum particularly produces cannabinoids bearing sesquiterpene moiety such as daurichromenic acid (DCA), grifolic acid (GFA), confluentin, and rhododaurichromenic acid [
11,
12,
13].
Rh. adamsii produces cannabigerorcinic acid, DCA, cannabigerorcinic acid methylase, chromane, and chromene monoterpenoids;
Rh. rubiginosum produces cannabinoid rubiginosins A–G [
14,
15].
Rh. anthopogonoides contains chromane/chromene derivatives such as cannabiorcicyclolic acid, cannabiorcichromenic acid, anthopogochromenic acid, and anthopogocyclolic acid (
Figure 3) [
16].
Figure 3. Structure of phytocannabinoids in Rhododendron plants.
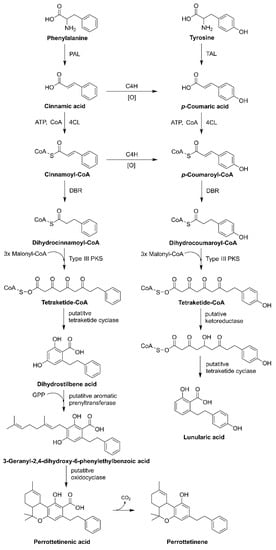
Liverworts, such as
Radula marginata,
R. perrottetii, and
R. laxirameae native to New Zealand, produce active cannabinoids with bibenzyl backbones such as lunularic acid and its dimeric form—vittatin (
Figure 4) [
17,
18,
19,
20]. Some fungi, e.g.,
Albatrellus (Albatrellaceae, mycorrhizal fungi) species, also produce GFA along with its derivative confluentin, grifolin, and neogrifolin (
Figure 4). Additionally,
Cylindrocarpon olidum generates cannabiorcichromenic acid and halogenated cannabinoid, i.e., 8-chlorocannabiorcichromenic acid (
Figure 4) [
1,
21].
Figure 4. Structure of phytocannabinoids in (a) liverworts and (b) fungi.
This review focuses on the biosynthesis of different active phytocannabinoids in several cellular compartments of C. sativa, Rhododendron, and Radula species. In this topic framework, the most crucial criterion is the synthetic and biotechnological techniques for the production of phytocannabinoids. The current review highlights the multi-faceted role of different active phytocannabinoids in humans and plants. Interestingly, this review briefly highlights the antimicrobial, antibacterial, and antibiotic properties of phytocannabinoid based on recent papers. Additionally, the role of phytocannabinoids in ameliorating pathogenic attack, and environmental stresses, e.g., cold, heat, and UV radiation, is also briefly assessed.
2. Phytocannabinoid Biosynthesis Sites
In
C. sativa, phytocannabinoids are stored in glandular trichomes, located all over the aerial part of the plant, so root surface and root tissues do not keep phytocannabinoid. Female flowers possess a high density of phytocannabinoid [
22,
23]. Glandular trichomes have balloon shaped secretory vessicle which store cannabinoid. High temperature or herbivory leads to trichome rupture, which releases the sticky contents on the plant parts with viscous and non-crystallizing properties [
24,
25]. Higher temperatures increase cannabinoid production. Furthermore, cannabinoid production is raised in the cannabis flower after UV-B exposure. Nevertheless, phytocannabinoids act as a sun shield that absorbs lethal UV radiation [
26].
Rhododendron genus lepidote consists of small leaves surrounded by glandular scales on both the abaxial and adaxial surfaces. These scales have lipophilic globules that contain major bioactive compounds such as cannabinoids, terpenes; the apoplastic space of the glandular scale also contains cannabinoids such as DCA in the
Rhododendron [
27]. Liverworts have oil bodies that are membrane-bound cellular structures that contain cannabinoids, aromatic oil, and terpenoid (
cis configuration), mostly sesquiterpenoids and diterpenoids. Oil bodies are odiferous bitter, pungent compounds, which make them biologically active. Furthermore, these possess several ecological advantages such as tolerance from temperature, light, or radiation [
28,
29].