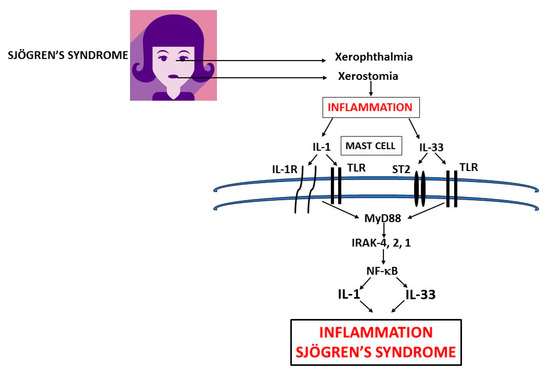
Sjögren's syndrome (SS) is a chronic autoimmune inflammatory disease which affects primarily older women and is characterized by irreversible damage of the exocrine glands, including tear (xerophthalmia) and salivary glands (xerostomia). Mast cells (MCs) are ubiquitous, but are primarily located close to blood vessels and nerves and can be activated early in autoimmune diseases, including SS, to express a wide variety of cytokines and chemokines. Activation of MCs by IL-1 and IL-33 in SS plays a crucial role and mediates the inflammatory process. Inhibitory effect of pro-inflammatory cytokines could be a new therapeutic strategy in the treatment of SS.
- Sjogren's Sindrome, Immunology, inflammation
1. Definition
Sjögren’s syndrome (SS) is a chronic autoimmune inflammatory disease that affects primarily older women and is characterized by irreversible damage of the exocrine glands, including tear (xerophthalmia) and salivary glands (xerostomia). Secretory glands lose their functionality due to the infiltration of immune cells, which produce cytokines and cause inflammation. Primary SS is characterized by dry syndrome with or without systemic commitment in the absence of other pathologies. Secondary SS is accompanied by other autoimmune diseases with high activation of B lymphocytes and the production of autoantibodies, including the rheumatoid factor. Other cells, such as CD4+ T cells and mast cells (MCs), participate in SS inflammation. MCs are ubiquitous, but are primarily located close to blood vessels and nerves and can be activated early in autoimmune diseases to express a wide variety of cytokines and chemokines. In the SS acute phase, MCs react by generating chemical mediators of inflammation, tumor necrosis factor (TNF), and other pro-inflammatory cytokines such as interleukin (IL)-1 and IL-33.
2. History
The first description of autoimmune diseases occurred in the late 19th century. Today, there are over 80 autoimmune disorders, where it is noted that the immune system attacks its cells or tissues, causing functional damage to the organs [1]. About 7% of Americans were revealed to have an autoimmune disease, of whom women are more commonly affected. Normally, autoimmune diseases arise during adulthood and the cause is generally unknown [2].
The causes of SS are unknown, but this is an autoimmune disease in which genetic factors and viral infections can play a role in the development of the pathogenesis which causes irreversible damage to the exocrine glands [4]. Individuals with pSS experience tear gland dysfunctions with xerophthalmia, where the eye cannot produce tears, the salivary glands are also damaged, causing dry mouth due to the lack of saliva (xerostomia), and other systemic symptoms [5]. The secretory glands lose their functionality due to the infiltration of immune cells, including lymphocytes, which release inflammatory cytokines, activate other immune cells, and cause inflammation [6] (Figure 1). SS can be classified as primary, characterized by dry syndrome with or without systemic commitment and in the absence of other pathologies, or secondary, which is accompanied by other autoimmune diseases, such as systemic lupus erythematosus, scleroderma, rheumatoid arthritis, Hashimoto’s thyroiditis [7] (in this paper we mainly deal with pSS, however, the inflammatory phenomenon can occur in both the primary and secondary form of SS). In pSS, there is high activation of B lymphocytes with the production of autoantibodies such as anti-Ro60 and anti-La/SSB [8]. In this disease, the immune state appears mediated by T cells and excessive activation of B lymphocytes with the generation of autoantibodies such as anti-Ro/60 and anti-La/SSB, directed towards nuclear antigens. In addition, in the more common secondary SS form, associated with other rheumatic autoimmune diseases, there is an infiltration of immune cells in various organs, such as endocrine glands, and deposition of various autoantibodies including rheumatoid factor (RF), anti-Ro60/SS-A and anti-La/SS-B, which provoke inflammation mediated by pro-inflammatory cytokines [9]. Antibodies to Ro60 antigens are called anti-SSA/Ro and patients who express Ro60 antibodies in most cases are diagnosed with autoimmune conditions, including SS. Ro/La antigens are made up of 52Ro, 60Ro and La proteins, in addition to RNA particles. Anti-Ro/60 is the most common form in autoimmune diseases and is also associated with SS. Auto-antibodies damage the exocrine glands and induce inflammation caused by cytokines and chemokines [10]. The pathological aspect is assumed to have multifactorial etiology where genetic, hormonal, and environmental factors may play a relevant role. Generally, pSS seems to be associated with a genetic predisposition linked to HLA-B (HLA-Dw3, HLA-DRw52 and HLA-DR3) and individuals who have this genetic picture have a 20-fold greater risk of developing the disease than others [11].
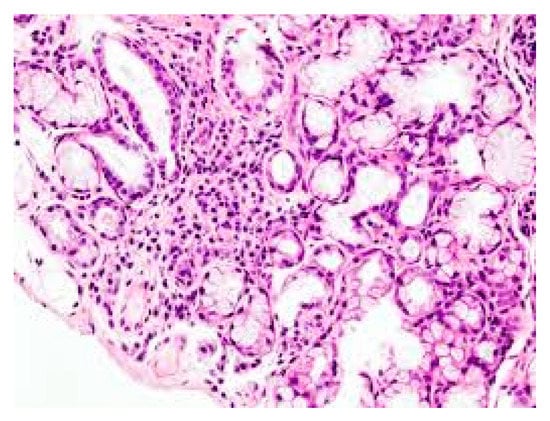
3. Data, Model, Applications and Influences
3.1. Mast Cells (MCs)
References
- Galli, S.J.; Kalesnikoff, J.; Grimbaldeston, M.A.; Piliponsky, A.M.; Williams, C.M.; Tsai, M. Mast cells as “tunable” effector and immunoregulatory cells: Recent advances. Annu. Rev. Immunol. 2005, 23, 749–786. [Google Scholar] [CrossRef]
- Kritas, S.K.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Conti, P. Mast cells contribute to coronavirus-induced inflammation: New anti-inflammatory strategy. J. Biol. Regu.l Homeost. Agents 2020, 34. [Google Scholar]
- Franza, L.; Carusi, V.; Altamura, S.; Caraffa, A.; E Gallenga, C.; Kritas, S.K.; Ronconi, G.; Conti, P.; Pandolfi, F. Interrelationship between inflammatory cytokines (IL-1, IL-6, IL-33, IL-37) and acquired immunity. J. Biol. Regul. Homeost. Agents 2019, 33, 1321–1326. [Google Scholar] [PubMed]
- Mukai, K.; Tsai, M.; Starkl, P.; Marichal, T.; Galli, S.J. IgE and mast cells in host defense against parasites and venoms. Semin. Immunopathol. 2016, 38, 581–603. [Google Scholar] [CrossRef]
- Galli, S.J.; Maurer, M.; Lantz, C.S. Mast cells as sentinels of innate immunity. Curr. Opin. Immunol. 1999, 11, 53–59. [Google Scholar] [CrossRef]
- Caraffa, A.; Gallenga, C.E.; Kritas, S.K.; Ronconi, G.; Conti, P. Impact of mast cells in systemic lupus erythematosus: Can inflammation be inhibited? J. Biol. Regul. Homeost. Agents 2019, 33, 669–673. [Google Scholar]
- Nakae, S.; Suto, H.; Berry, G.J.; Galli, S.J. Mast cell–derived TNF can promote Th17 cell–dependent neutrophil recruitment in ovalbumin-challenged OTII mice. Blood 2006, 109, 3640–3648. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Lessiani, G.; Kritas, S.K.; Ronconi, G.; Caraffa, A.L.; Theoharides, T.C. Mast cells emerge as mediators of atherosclerosis: Special emphasis on IL-37 inhibition. Tissue Cell 2017, 49, 393–400. [Google Scholar] [CrossRef]
- Conti, P.; Lauritano, D.; Caraffa, A.; Gallenga, C.E.; Kritas, S.K.; Ronconi, G.; Martinotti, S. Microglia and mast cells generate proinflammatory cytokines in the brain and worsen inflammatory state: Suppressor effect of IL-37. Eur. J. Pharmacol. 2020, 875, 173035. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Nakae, S.; Tsai, M. Mast cells in the development of adaptive immune responses. Nat. Immunol. 2005, 6, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.-L.; Gao, J.-M.; Li, P.-P.; Wang, X. IL-9 Contributes to Immunosuppression Mediated by Regulatory T Cells and Mast Cells in B-Cell Non-Hodgkin’s Lymphoma. J. Clin. Immunol. 2011, 31, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Chavarría, A.; Alcocer-Varela, J. Is damage in central nervous system due to inflammation? Autoimmun. Rev. 2004, 3, 251–260. [Google Scholar] [CrossRef]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Brown, M.A.; Hural, J. Functions of IL-4 and Control of Its Expression. Crit. Rev. Immunol. 2017, 37, 181–212. [Google Scholar] [CrossRef]
- Svenningsen, S.; Nair, P. Asthma Endotypes and an Overview of Targeted Therapy for Asthma. Front. Med. 2017, 4, 158. [Google Scholar] [CrossRef]
- Liu, X.; Xiao, Y.; Pan, Y.; He, L.; Zheng, S.G.; Wenru, S. The role of the IL-33/ST2 axis in autoimmune disorders: Friend or foe? Cytokine Growth Factor Rev. 2019, 50, 60–74. [Google Scholar] [CrossRef]
- Lauritano, D.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Kritas, S.K.; di Emidio, P.; Martinotti, S.; Tetè, G.; Ross, R.; Conti, P. New aspect of allergic contact dermatitis, an inflammatory skin disorder mediated by mast cells: Can IL-38 help? Med. Hypotheses 2020, 139, 109687. [Google Scholar] [CrossRef] [PubMed]
- Shaik, Y.; Caraffa, A.; Ronconi, G.; Lessiani, G.; Conti, P. Impact of polyphenols on mast cells with special emphasis on the effect of quercetin and luteolin. Central Eur. J. Immunol. 2018, 43, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.I.; Lee, K.H.; Joo, Y.H.; Lee, J.M.; Jeon, J.; Jung, H.J.; Shin, M.; Cho, S.; Kim, T.H.; Park, S.; et al. Inflammasomes and autoimmune and rheumatic diseases: A comprehensive review. J. Autoimmun. 2019, 103, 102299. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhang, X.; Wang, M.; Chen, Z.; Yan, Y.; Gu, W.; Tan, J.; Jiang, W.; Ji, W. Exosomes from Thymic Stromal Lymphopoietin-Activated Dendritic Cells Promote Th2 Differentiation through the OX40 Ligand. Pathobiology 2018, 86, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Andonegui, G.; Wong, C.H.; Kubes, P. Role of endothelial TLR4 for neutrophil recruitment into central nervous system microvessels in systemic inflammation. J. Immunol. 2009, 183, 5244–5250. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Kalesnikoff, J.; Grimbaldeston, M.A.; Piliponsky, A.M.; Williams, C.M.; Tsai, M. Mast cells as “tunable” effector and immunoregulatory cells: Recent advances. Annu. Rev. Immunol. 2005, 23, 749–786. [Google Scholar] [CrossRef]
- Kritas, S.K.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Conti, P. Mast cells contribute to coronavirus-induced inflammation: New anti-inflammatory strategy. J. Biol. Regu.l Homeost. Agents 2020, 34. [Google Scholar]
- Franza, L.; Carusi, V.; Altamura, S.; Caraffa, A.; E Gallenga, C.; Kritas, S.K.; Ronconi, G.; Conti, P.; Pandolfi, F. Interrelationship between inflammatory cytokines (IL-1, IL-6, IL-33, IL-37) and acquired immunity. J. Biol. Regul. Homeost. Agents 2019, 33, 1321–1326. [Google Scholar] [PubMed]
- Mukai, K.; Tsai, M.; Starkl, P.; Marichal, T.; Galli, S.J. IgE and mast cells in host defense against parasites and venoms. Semin. Immunopathol. 2016, 38, 581–603. [Google Scholar] [CrossRef]
- Galli, S.J.; Maurer, M.; Lantz, C.S. Mast cells as sentinels of innate immunity. Curr. Opin. Immunol. 1999, 11, 53–59. [Google Scholar] [CrossRef]
- Caraffa, A.; Gallenga, C.E.; Kritas, S.K.; Ronconi, G.; Conti, P. Impact of mast cells in systemic lupus erythematosus: Can inflammation be inhibited? J. Biol. Regul. Homeost. Agents 2019, 33, 669–673. [Google Scholar]
- Nakae, S.; Suto, H.; Berry, G.J.; Galli, S.J. Mast cell–derived TNF can promote Th17 cell–dependent neutrophil recruitment in ovalbumin-challenged OTII mice. Blood 2006, 109, 3640–3648. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Lessiani, G.; Kritas, S.K.; Ronconi, G.; Caraffa, A.L.; Theoharides, T.C. Mast cells emerge as mediators of atherosclerosis: Special emphasis on IL-37 inhibition. Tissue Cell 2017, 49, 393–400. [Google Scholar] [CrossRef]
- Conti, P.; Lauritano, D.; Caraffa, A.; Gallenga, C.E.; Kritas, S.K.; Ronconi, G.; Martinotti, S. Microglia and mast cells generate proinflammatory cytokines in the brain and worsen inflammatory state: Suppressor effect of IL-37. Eur. J. Pharmacol. 2020, 875, 173035. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Nakae, S.; Tsai, M. Mast cells in the development of adaptive immune responses. Nat. Immunol. 2005, 6, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.-L.; Gao, J.-M.; Li, P.-P.; Wang, X. IL-9 Contributes to Immunosuppression Mediated by Regulatory T Cells and Mast Cells in B-Cell Non-Hodgkin’s Lymphoma. J. Clin. Immunol. 2011, 31, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Chavarría, A.; Alcocer-Varela, J. Is damage in central nervous system due to inflammation? Autoimmun. Rev. 2004, 3, 251–260. [Google Scholar] [CrossRef]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Brown, M.A.; Hural, J. Functions of IL-4 and Control of Its Expression. Crit. Rev. Immunol. 2017, 37, 181–212. [Google Scholar] [CrossRef]
- Svenningsen, S.; Nair, P. Asthma Endotypes and an Overview of Targeted Therapy for Asthma. Front. Med. 2017, 4, 158. [Google Scholar] [CrossRef]
- Liu, X.; Xiao, Y.; Pan, Y.; He, L.; Zheng, S.G.; Wenru, S. The role of the IL-33/ST2 axis in autoimmune disorders: Friend or foe? Cytokine Growth Factor Rev. 2019, 50, 60–74. [Google Scholar] [CrossRef]
- Lauritano, D.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Kritas, S.K.; di Emidio, P.; Martinotti, S.; Tetè, G.; Ross, R.; Conti, P. New aspect of allergic contact dermatitis, an inflammatory skin disorder mediated by mast cells: Can IL-38 help? Med. Hypotheses 2020, 139, 109687. [Google Scholar] [CrossRef] [PubMed]
- Shaik, Y.; Caraffa, A.; Ronconi, G.; Lessiani, G.; Conti, P. Impact of polyphenols on mast cells with special emphasis on the effect of quercetin and luteolin. Central Eur. J. Immunol. 2018, 43, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.I.; Lee, K.H.; Joo, Y.H.; Lee, J.M.; Jeon, J.; Jung, H.J.; Shin, M.; Cho, S.; Kim, T.H.; Park, S.; et al. Inflammasomes and autoimmune and rheumatic diseases: A comprehensive review. J. Autoimmun. 2019, 103, 102299. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhang, X.; Wang, M.; Chen, Z.; Yan, Y.; Gu, W.; Tan, J.; Jiang, W.; Ji, W. Exosomes from Thymic Stromal Lymphopoietin-Activated Dendritic Cells Promote Th2 Differentiation through the OX40 Ligand. Pathobiology 2018, 86, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Andonegui, G.; Wong, C.H.; Kubes, P. Role of endothelial TLR4 for neutrophil recruitment into central nervous system microvessels in systemic inflammation. J. Immunol. 2009, 183, 5244–5250. [Google Scholar] [CrossRef] [PubMed]
This entry is adapted from the peer-reviewed paper 10.3390/ijms21124297