Fibroblastic reticular cells (FRCs), usually found and isolated from the T cell zone of lymph nodes, have recently been described as much more than simple structural cells. Originally, these cells were described to form a conduit system called the “reticular fiber network” and for being responsible for transferring the lymph fluid drained from tissues through afferent lymphatic vessels to the T cell zone. However, nowadays, these cells are described as being capable of secreting several cytokines and chemokines and possessing the ability to interfere with the immune response, improving it, and also controlling lymphocyte proliferation. Here, we performed a systematic review of the several methods employed to investigate the mechanisms used by fibroblastic reticular cells to control the immune response, as well as their ability in determining the fate of T cells. We searched articles indexed and published in the last five years, between 2016 and 2020, in PubMed, Scopus, and Cochrane, following the PRISMA guidelines. We found 175 articles published in the literature using our searching strategies, but only 24 articles fulfilled our inclusion criteria and are discussed here. Other articles important in the built knowledge of FRCs were included in the introduction and discussion. The studies selected for this review used different strategies in order to access the contribution of FRCs to different mechanisms involved in the immune response: 21% evaluated viral infection in this context, 13% used a model of autoimmunity, 8% used a model of GvHD or cancer, 4% used a model of Ischemic-reperfusion injury (IRI). Another four studies just targeted a particular signaling pathway, such as MHC II expression, FRC microvesicles, FRC secretion of IL-15, FRC network, or ablation of the lysophosphatidic acid (LPA)-producing ectoenzyme autotaxin. In conclusion, our review shows the strategies used by several studies to isolate and culture fibroblastic reticular cells, the models chosen by each one, and dissects their main findings and implications in homeostasis and disease.
Lately, FRCs’ ability for controlling the immune response and its role in several pathological conditions, such as viral infection, inflammation, metastatic cancer, and autoimmunity, are also included in this review. Consequently, we comprise here the latest updates in FRC biology, their impact on T cell fate, how they participate in diseases, and how they could be manipulated in order to ameliorate the course of certain conditions.
FRC is a specific subset of stromal cells present in the lymph node, and they are precisely located in the T cell zone. There are other stromal cell subsets in lymph nodes, described as double-negative cells, follicular dendritic cells, blood endothelial cells, lymphatic endothelial cells, and others that are not discussed in this review
[42].
The results of this review firstly show the characteristics of the host type used for analyzing FRC function. In addition, strategies used by them in order to achieve their target objectives, including model characteristics, such as source, genotype, age, and gender, are described in . The main characteristics of FRCs, their origin, as well the lymph node (LN) digestion process, and techniques used for their isolation are described in . The immune cell sources, as well as their characteristics, are described in . All these variations between the models studied, cell origins, and characterization, sometimes lead to different conclusions, making the comparison between studies difficult or conflicting, such as the role of FRCs in T cell proliferation, sometimes described as stimulators and, at other times, as limiting. Next, we assembled the studies with the same subject () and compared them, trying to show the differences and, more importantly, comparisons between the achieved results ().
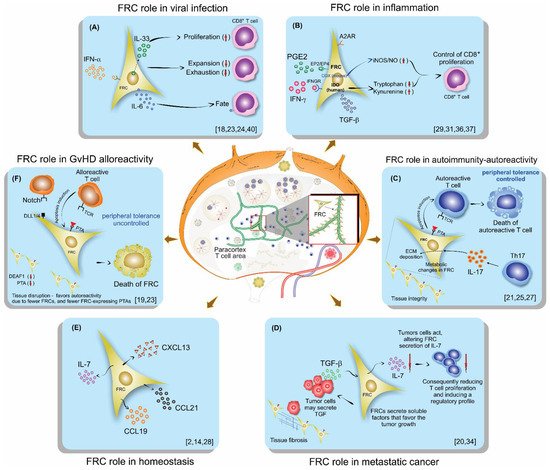
Figure 1. Schematic illustration of lymph nodes, FRC localization, and their role on lymphocytes in different scenarios of the immune response: (A) Viral infection, (B) Inflammation, (C) Autoimmunity, (D) Metastatic cancer, (E) Homeostasis, (F) GvHD.
The first scenario discussed was on viral infection (A). Aparicio-Domingo et al., in an LCMV study, concluded that FRCs displayed a stimulatory role, being a main source of IL-33 in the lymph node and crucial for leading to acute and chronic antiviral T cell responses. They also showed that FRCs mainly act on CD8 T lymphocytes by signaling via ST2 expressed by these T cells
[18]. Severino et al. demonstrated previously, in 2017, the increased IL-33 gene expression in human FRCs after treatment with IFN-γ or IL-1β and TNF-α. These cytokines are usually released during a course of an immune response, supporting the Aparicio-Domingo et al. findings that FRCs are the main source for IL-33
[9].
Perez-Shibayama et al., using the LCMV model like Aparicio-Domingo et al., commented that FRCs contributed to an immunostimulatory state to prevent virus replication and spread. However, they also found a regulatory role of FRCs, showing an IFN-α-signaling dependent shift of FRCs toward an immunoregulatory state, reducing exhaustive CD8 T lymphocyte activation. They claim that type 1 IFN-mediated control of LCMV replication in FRCs is one of the major factors that determine the quality of the antiviral CD8+ T cell response
[23]. In agreement, Talemi and Hofer sustain the idea that interferons delay the viral spread in infection, acting as sentinels, warning uninfected cells, and also are negative feedback regulators acting at a single-cell level
[43].
Regarding the anti-viral response for influenza and LCMV, Brown et al.
[24] showed that FRCs function is more than controlling T cell expansion. FRCs also outline the fate and function of CD8 T lymphocytes through their IL-6 production, and CD8 T cells exposed to both FCRs and IL-6 are driven to a memory phenotype. In addition, CD8 T cells cultivated in the presence of FRCs are more persistent during a viral infection than CD8 T cells stimulated without FRC presence
[24]. Moreover, the pleiotropic function and the importance of IL-6 were reported before, supporting that this cytokine, in certain environments, could be an important player for guiding the immune response
[44]. Next, Masters et al., reported that after aging-related changes, FRCs have an altered impact on the beginning of the immune response to influenza infection, consequently contributing to delayed T lymphocytes responses to this virus
[28]. Moreover, their findings on the importance of homeostatic chemokines for the success of the anti-viral response are also supported by Chai et al., who previously reported on the importance of these chemokines secreted by FRCs to the immune response against virus infection, and by Thompson et al., who also reported on the role of the lymph node in aging mice and its negative impact on T cells
[45][46]. Lastly, Royer et al. proposed that HSV-1 in lymph nodes can cause pathological alterations in the FRC conduit system, resulting in fewer HSV-specific CD8 T lymphocytes in circulation, and a diminished anti-viral response to this virus. In addition, they claim that immunodeficiency can occur as a secondary outcome of FRC alterations to SLOs
[40]. Their results are supported by other models that impair T cell responses due to virus-associated damage to FRCs
[47][48].
Concerning inflammation (B) and the COX/PGE
2 pathway, which converts arachidonic acid in several prostanoids via the enzymes COX1 and COX2, FRCs have been proposed to play dual roles by either promoting or inhibiting adaptive immunity
[49][50], similar to myeloid and T cells. Schaeuble et al.’s experiments revealed that FRCs can control T cell responses, independently of other cells, by two pathways that lead to NO release, clarifying that one pathway is activated via the sensing of IFN-y by FRCs, which is activated only by strong T cell responses, and another pathway is mediated by COX2-dependent synthesis of PGE
2, which signals via EP1 and EP2 during both weak and strong T cells responses
[29]. Knoblich et al. also demonstrated that FRCs control T cell proliferation and modulate their differentiation
[31]. Knoblich et al. included even more mechanisms that control T cell proliferation besides IFN-y and PGE
2, which, in human cells, do not release NO, but instead activate IDO; they point to TGF-β and the adenosine 2A receptor (A2AR) as other signaling pathways affecting T cell proliferation. They also demonstrated that human FRCs affect the fate of naïve T cells, diminishing their differentiation into central memory while enhancing effector and effector memory phenotypes
[31]. Yu, M. et al. support these findings with their previous study on the animal model and in vitro assays, confirming that hyperactive COX-2/PGE
2 pathways in FRCs are a mechanism that maintains peripheral T cell tolerance
[37]. In addition, Valencia et al. demonstrated the differences between mice and humans regarding COX inflammatory pathways, and concluded that human and murine FRCs would regulate T lymphocytes responses using different mechanisms, and arguing that, in humans, IDO would play a more important role than iNOS/NO
[36].
Further, in autoimmunity, the FRC network seems to play an important role (C). Gonzalez et al., using a type 1 diabetes (T1D) model and a 3D system of culture, found that in T1D FRCs, the reticular network organization was altered, displayed larger pores, and had a lower expression of podoplanin compared to a control animal or control culture system. They also demonstrated a reduced expression of PTAs and T1D antigens in T1D FRCs. Consequently, FRCs modulated their interactions with autoreactive T lymphocytes by remodeling their reticular network in LNs; PTAs and podoplanin played a central role and their alterations may favor T1D
[21]. These findings are supported by a previous study from the same group that investigated alterations in pancreatic lymph nodes from humans and mice
[51]. Kasinath et al. studied crescentic glomerulonephritis (GN), an autoimmune inflammatory condition characterized by the rapid deterioration of kidney function. They investigated the role of fibroblastic reticular cells residing in the stromal compartment of the kidney lymph node in this model. They observed that FRCs are fundamental to the propagation of the immune response in nephrotoxic serum nephritis. Following GN development, they observed an increase in effector memory and Th17 cells in the kidney LN. In addition, they observed that the removal of the kidney lymph node, a depletion of fibroblastic reticular cells, and treatment with anti-podoplanin antibodies each resulted in a reduction of kidney injury
[25]. Majumder et al. studied the EAE model, and they also showed Th17 differentiation in LNs and that the signaling in the receptor for IL-17 in FRCs is related to collagen deposition in LNs. This work suggests that Th17 cells promote ECM deposition in inflamed LNs through FRCs-IL-17 signaling, independently of LN size or hypercellularity. As a consequence of Th17 in LN, the released IL-17 signals in FRCs impact LN stromal organization, leading to FRC activation by changing their phenotype from quiescence to highly metabolic. Moreover, the absence of IL-17 signaling in FRCs does not lead to immune failings but does cause impaired B cell responses, due to the reduced availability of BAFF, which is critical for the germinal center formation and maintenance
[27].
In metastatic cancer (D), FRCs appear to be regulated by the tumor cells. Eom et al. showed in human melanoma that FRCs in tumor-infiltrated LNs may favor cancer invasion and progression through secretion of soluble factors, alterations in the lymph node structure, and by promoting pathological conditions such as fibrosis
[20]. Gao et al. also showed in tumor-infiltrated LNs a decrease in FRCs and IL-7 secretion, leading to a declined number and diminished function of T cells in LNs
[34].
In homeostasis, as displayed in E, FRCs played an important role in secreting homeostatic chemokines, promoting the meeting between T cells and dendritic cells on the T cell zone, and also by secreting IL-7, an essential cytokine involved in T cell effector memory differentiation
[2][14][28].
Furthermore, in graft versus host disease (GvHD) FRCs’ ability for peripheral tissue antigen (PTA) presentation and NOTCH signaling have been shown to be important features in the aggravation and maintenance of the GvHD state (F). Dertschnig et al. showed that the loss of PTA presentation by FRCs during GVHD leads to permanent damage in their networks in lymphoid tissues, compromising peripheral tolerance. In addition, they demonstrated that not only the disruption of FCRs occurs during GvHD but also the capacity for the regeneration of this network is impaired, different to what was found for viral infection, where the damage occurs, but after viral clearance, the network is restored
[19]. Chung et al. showed that FRC-delivered NOTCH signals through the ligands DLL1 and DLL4 to donor alloreactive T cells help to program the pathogenicity of these T cells. Moreover, they demonstrated that the early use of neutralizing antibodies against DDL1 and DDL4 abrogated GvHD
[33].
As seen by Kasinath et al. in GN
[25], Maaraouf et al., using ischemic reperfusion injury (IRI) with multiple IRI
[32], reported that kidney LNs (KLNs) cause excessive deposition of ECM fibers containing fibronectin and collagen, which leads to local fibrosis, similar to kidney fibrosis. They confirmed that depletion of FRCs reduced T cell activation in the KLNs and ameliorated renal injury in acute IRI
[25][32].
Regarding pathway investigation, Knop et al. demonstrated that FRC-derived IL-7 plays an essential role in maintaining central memory T cells, but is dispensable for naïve T cell survival
[22]. Dubrot et al. showed a mechanism of T lymphocyte proliferation inhibition by the IFN-y-induced expression of MHC II
[30]. In addition, they demonstrated that the deletion of MHC II in LN stromal cells in vivo leads to diminished Treg frequency and functions, and, at the same time, enhances effector cell differentiation, further leading to T cell tissue infiltration and the subsequent development of T cell-mediated autoimmunity
[30]. Pazstoi et al. used the stromal compartment of gut-draining LNs to demonstrate that FRCs own the tolerogenic capacity that controls T cells. They also demonstrated that mesenteric LNs (mLNs) are more capable of inducing
[35] Treg profiles than the peripheral ones. Likewise, they demonstrated that FRCs contribute to peripheral tolerance by developing de novo Treg by releasing microvesicles (MVEs), which carry high levels of TGF-β
[35]. Gil-Cruz et al. also used mLNs and Peyer’s patches (PP) as the targets of their study and identified that an antiviral response driven by ILC1 and NK was regulated by the FRCs’ limiting provision of IL-15
[38]. This mechanism control seems to be activated by TLR7 and/or IL-1β, and its control is designated by the MyD88-dependent pathway
[38]. Novkovic confirmed that the physical scaffold of LNs formed by the FRC network is critical for the maintenance and functionality of LNs
[39], and Takeda et al. demonstrated the role of LPA derived from FRCs in T cell motility
[41]. Kelch et al. demonstrated, by imaging, the conduit density in the deep and superficial T cell zone, concluding that although T cells within the superficial zone stay in constant contact with FRCs, and in the deep T cell zone, there is a gap that does not guarantee simultaneous contact for all T cells in this region
[26].
In summary, FRCs in homeostasis plays an important role in secreting homeostatic chemokines and IL-7, which are essential for the immune response initiation and for T cell effector memory phenotype differentiation. In a viral setting, FRCs are the main source of IL-33, playing a regulatory role by diminishing the T cell exhaustion, and acting on T cell fate through IL-6 secretion. In this same setting, aging FRCs have a negative impact on T cells. In inflammation, FRCs have been proposed to play a dual role by either promoting or inhibiting adaptive immunity. The main mechanisms behind inflammation are related to IFN-y and PGE2-signaling that, in murine cells, release NO and, in humans, activate IDO. In autoimmunity, the reticular network organization was altered, displayed lower expression of PTAs and podoplanin, and, in this context, IL-17 signaling impacted LN stromal organization, leading to highly metabolically activated FRCs. In metastatic cancer, FRCs appear to be regulated by the tumor cells decreasing IL-7 secretion and enhancing other soluble factors, causing alterations in the lymph node structure, such as fibrosis. In GvHD, the loss of PTA presentation by FRCs leads to permanent damage in their networks, compromising peripheral tolerance.