The oral cavity is a portal into the digestive system, which exhibits unique sensory properties. Like facial skin, the oral mucosa needs to be exquisitely sensitive and selective, in order to detect harmful toxins versus edible food. Chemosensation and somatosensation by multiple receptors, including transient receptor potential channels, are well-developed to meet these needs. In contrast to facial skin, however, the oral mucosa rarely exhibits itch responses. Like the gut, the oral cavity performs mechanical and chemical digestion. Therefore, the oral mucosa needs to be insensitive, to some degree, in order to endure noxious irritation. Persistent pain from the oral mucosa is often due to ulcers, involving both tissue injury and infection. Trigeminal nerve injury and trigeminal neuralgia produce intractable pain in the orofacial skin and the oral mucosa, through mechanisms distinct from those seen in the spinal area, which is particularly difficult to predict or treat. The diagnosis and treatment of idiopathic chronic pain, such as atypical odontalgia (idiopathic painful trigeminal neuropathy or post-traumatic trigeminal neuropathy) and burning mouth syndrome, remain especially challenging. The central integration of gustatory inputs might modulate chronic oral and facial pain. A lack of pain in chronic inflammation inside the oral cavity, such as chronic periodontitis, involves the specialized functioning of oral bacteria. A more detailed understanding of the unique neurobiology of pain from the orofacial skin and the oral mucosa should help us develop novel methods for better treating persistent orofacial pain.
- chronic pain
- mucosa pain
- orofacial pain
1. Introduction
| Table | Subtype |
|---|---|
| 1. Orofacial pain attributed to disorders of dentoalveolar and anatomically related structures | 1.1 Dental pain 1.2 Oral mucosal, salivary gland, and jawbone pains |
| 2. Myofascial orofacial pain | 2.1 Primary myofascial orofacial pain 2.2 Secondary myofascial orofacial pain |
| 3. Temporomandibular joint (TMJ) pain | 3.1 Primary temporomandibular joint pain 3.2 Secondary temporomandibular joint pain |
| 4. Orofacial pain attributed to lesion or disease of the cranial nerves | 4.1Pain attributed to lesion or diseaseofthetrigeminalnerve 4.2 Pain attributed to lesion or disease of the glossopharyngeal nerve |
| 5. Orofacial pains resembling presentations of primary headaches | 5.1 Orofacial migraine 5.2 Tension-type orofacial pain 5.3 Trigeminal autonomic orofacial pain 5.4 Neurovascular orofacial pain |
| 6. Idiopathic orofacial pain | 6.1 Burning mouth syndrome (BMS) 6.2 Persistent idiopathic facial pain (PIFP) 6.3 Persistent idiopathic dentoalveolar pain 6.4 Constant unilateral facial pain with additional attacks (CUFPA) |
2. Physiological Somatosensation and Pain from Oral Mucosa and Facial Skin
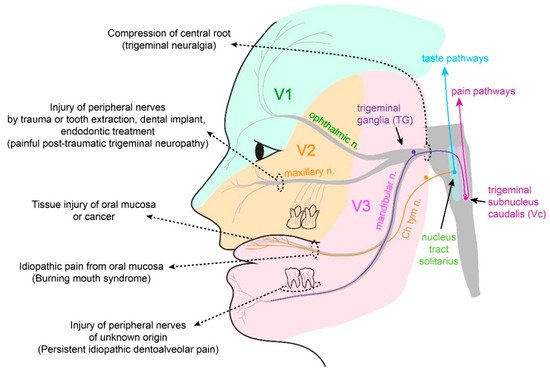
2.1. Somatosensation of Oral Mucosa and Facial Skin
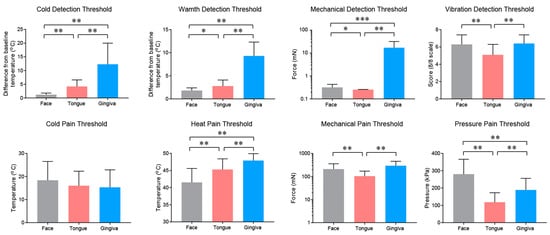
2.2. Properties and Projections of Primary Afferents in Oral Mucosa and Facial Skin
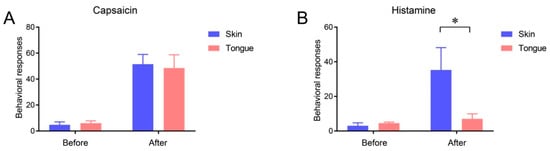
2.3. Itch Sensation of Oral Mucosa Is Weaker Than That of Facial Skin
This entry is adapted from the peer-reviewed paper 10.3390/ijms22115810