Biliary tract cancer (BTC) is often refractory to conventional therapeutics and is difficult to diagnose in the early stages. Implantation-based models have recently drawn attention for their convenience, flexibility, and scalability.
1. Introduction
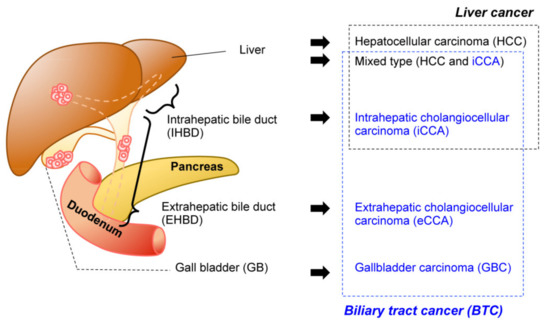
The biliary system is a network of bile ducts that collect bile produced and secreted by hepatocytes in the liver. The bile ducts merge into the common bile duct (CBD), transporting bile to the duodenum, where it aids the absorption of dietary lipids in the intestine. The gall bladder (GB) is located in the middle of this network. It functions to temporally store bile during the fast state, which in turn is secreted by GB contraction upon food intake. Biliary tract cancer (BTC) is a malignant tumorous cancer that arises from epithelial cells that cover the lumen of the bile duct. It is typically divided into three subtypes () based on anatomical site: intrahepatic cholangiocarcinoma (iCCA), extrahepatic cholangiocarcinoma (eCCA), and gallbladder carcinoma (GBC). While the histological diagnosis for most BTCs is adenocarcinoma, a subset of liver cancer is a biphasic tumor comprising both iCCA and hepatocellular carcinoma (HCC), thereby diagnosed as mixed type [
1].
Figure 1. Malignancy of the hepatobiliary system. Anatomical classification of the biliary system (left). The corresponding tumors are listed (right). Biliary tract cancers (BTC) (blue) and liver cancer (black) are not necessarily mutually exclusive.
The major risk factors for BTC include chronic infection with liver fluke and primary sclerosing cholangitis. Other risk factors include chronic liver disease, stones, fibrocystic polycystic disease, chemicals, obesity, aging, and some genetic diseases, suggesting that chronic inflammation in the local biliary tract may be implicated in its pathogenesis [
2]. Worldwide, iCCA cases are increasing, and eCCA cases are decreasing. The reasons for this trend remain largely unknown [
3,
4]. BTC is one of the most devastating cancer types, with a 5-year survival rate of approximately 10% to 30% for all patients and 2% to 3% for patients with metastatic disease. Although biliary intraepithelial neoplasia (BilIN) [
5,
6] is regarded as a putative pre-cancerous lesion for BTC, it can only be detected by histological examination of the tumors. Non-invasive diagnostic modalities, such as serum biomarkers or imaging, have not been developed. For early stage tumors, surgical resection followed by adjuvant chemotherapy is the first-line therapy to achieve a complete cure. In contrast, patients with advanced or metastatic disease are treated with systemic chemotherapy, radiation therapy, and local therapy, which may lead to palliative care.
Recent genomic analyses have revealed that mutations in tumor protein 53 (TP53), BReast CAncer gene (BRCA), phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha (PIK3CA), and KRAS are commonly found in all types of BTC. In contrast, fibroblast growth factor receptor 2 (FGFR2) fusion genes and isocitrate dehydrogenase 1 (IDH1) mutations are preferentially detected in iCCA, protein kinase CAMP-activated catalytic subunit alpha (PRKACA)- and PRKACB-fusion genes in eCCA, and mutations in epidermal growth factor receptor (EGFR), ERBB3, and phosphatase and tensin homolog (PTEN) in GBC [
7,
8,
9,
10,
11,
12,
13,
14]. Hence, molecular targeted therapies with inhibitors of IDH or FGFR can be considered for patients with iCCA. Radiation therapy and immunotherapy with immune checkpoint inhibitors or chimeric antigen receptor T cell therapy may also be applied, although their efficacy remains elusive. Mouse models, particularly genetically engineered mice (GEM), have been fundamentally important in elucidating the mechanisms underlying BTC development and are a potentially powerful tool for preclinical studies.
Given the recent progress in cancer genome projects and their application to genomic medicine in clinical practice, this work comprehensively reviews the recent progress in mouse BTC models, emphasizing those that recapitulate the whole processes of BTC development by genetic approaches. Although perihilar cholangiocarcinoma (pCCA) is a distinct entity of high clinical relevance, it is not distinguished from eCCA in most genomic studies, and no mouse model has been developed for pCCA. Therefore, we adopted this classification of BTC in this review and excluded hepatocellular carcinoma (HCC) models unless the induced tumors were mixed with iCCA. Similarly, other BTC models [
15,
16,
17] with patient-derived xenografts in immunodeficient mice, or in vivo mouse models based on the liver damage caused by chemicals or bile duct ligation were also excluded.
2. Technical Overview of Genetic Mouse Models of BTC
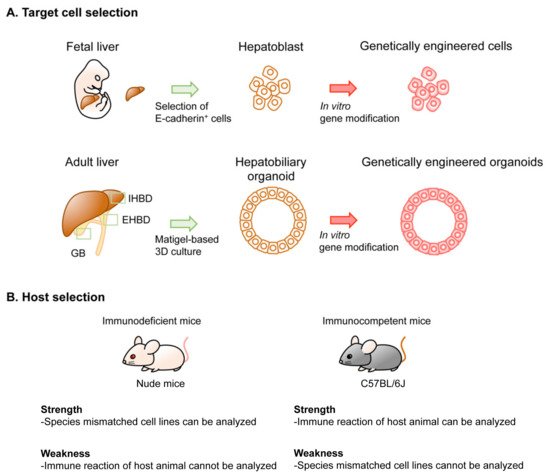
There are several different platforms for genetic modeling of BTC in mice, generating considerable variations in the latency and penetrance of tumor development, cost, equipment, technique required for the models, and the experimental settings. Thus, thorough considerations are advisable before selecting the type of model to be generated. Several options can be selected for the genetic modeling of BTC in mice. These include the target cell type, the method used in genetic engineering, the type of host mouse, and the location of tumor development (). In this section, we review the technical aspects of these options.
Figure 2. Options in implantation-based modeling of BTC. (See for the details of each study.) As examples of options for modeling BTC, target cell selection (A) and host selection (B) are illustrated. Abbreviations are: IHBD, intrahepatic bile duct and EHBD, extrahepatic bile duct. Nude mice and C57BL/6J strain mice were used as representatives for immunodeficient mice and immunocompetent mice, respectively.
Table 1. Implantation- and organoid-based hybrid model of biliary tract cancer in mice.
|
Driver
Oncogenes
|
Genotype of Organoids *
|
Methods for Genetic
Engineering **
|
Host
|
Implantation
|
Ref.
|
| |
|
Oncogenes
|
TSGs
|
|
|
|
|
HCC (Hepatocellular Carcinoma) from Liver Organoid
|
|
cMyc
|
WT
|
cDNA (R)
|
shRNA (R); Trp53 and CRISPR/Cas9 (T); Apc
|
C57BL/6J
|
liver
|
[18]
|
|
iCCA (Intrahepatic Cholangiocelular Carcinoma) from Liver Organoid
|
|
KrasG12D
|
KrasLSL-G12D/+
|
Cre (L)
|
shRNA (L): Cdkn2a and/or Pten, Trp53, Apc
|
Nude
|
s.c.
|
[19]
|
|
KrasLSL-G12D/+; Trp53flox/flox
|
N.T.
|
|
Pik3caH1047R
|
Pik3caH1047R
|
shRNA (L): Cdkn2a, Pten,
|
|
Rosa26- Pik3caH1047R; Trp53flox/flox
|
N.T.
|
|
FGFR2-AHCYL1
|
WT
|
cDNA(R)
|
shRNA (L): Cdkn2a and/or Pten
|
|
KrasG12D
|
KrasLSL-G12D/+; Trp53flox/flox (outbred)
|
Cre (R)
|
N.T. or shRNA (R): Pten
|
NSG
|
s.c. or liver
|
[18]
|
|
KrasLSL-G12D/+
|
Cre (T)
|
CRISPR/Cas9 (T): Pten and Trp53
|
C57BL/6J
|
liver
|
|
FGFR2-BICC1, -MGEA5, -TACC3
|
Trp53−/−
|
cDNA (R)
|
N.T.
|
NOD-SCID
|
s.c. or liver
|
[20]
|
|
KRASG12V
|
Cdkn2a−/−
|
cDNA (R)
|
|
C57BL/6J
|
s.c., liver, or kidney
|
[21]
|
|
eCCA (Extrahepatic Cholangiocelular Carcinoma) from CBD Organoid
|
|
KrasG12D
|
KrasLSL-G12D; Tgfbr2flox/flox; Cdh1flox/flox
|
Cre (L)
|
N.T.
|
Nude or C57BL/6J
|
s.c.
|
[22]
|
|
KRASG12V
|
Cdkn2a−/−
|
cDNA (R)
|
N.T.
|
C57BL/6J
|
s.c., liver, or kidney
|
[21]
|
|
GBC (Gallbladder Carcinoma) from GB Organoid
|
|
KrasG12D
|
KrasLSL-G12D
|
Cre (L)
|
shRNA (L): Cdkn2a, Pten
|
Nude
|
s.c.
|
[19]
|
|
KrasLSL-G12D/+; Trp53flox/flox
|
N.T.
|
|
KRASG12V
|
Cdkn2a−/−
|
cDNA (R)
|
N.T.
|
C57BL/6J
|
s.c., liver, or kidney
|
[21]
|
|
KrasG12D
|
WT
|
Cre (T)
|
CRISPR/Cas9 (T): Pten and Trp53
|
C57BL/6J
|
s.c. or GB
|
[23]
|
|
ERBB2S310F or V777L
|
cDNA (R)
|
CRISPR/Cas9 (T): Trp53
|
NSG
|
s.c.
|
| |
Rosa26- Pik3caH1047R; Trp53flox/flox
|
Cre (L)
|
N.T.
|
Nude
|
s.c.
|
[24]
|
|
KrasG12D
|
KrasLSL-G12D
|
CRISPR/Cas9 (T): Trp53, p19Arf, Smad4
|
C57BL/6J
|
GB via s.c.
|
For those models that developed cancer, pre-cancerous lesion and cystic lesions were listed. Note that liver organoid-derived tumor are either HCC or CCA (adenocarcinoma), but not mixed type liver cancer * C57BL/6J background unless otherwise indicated. ** L, lentivirus; R, retrovirus; T, transfection; N.T., not tested. Abbreviations: TSG, tumor suppressor genes; s.c., subcutis; WT, wildtype; CBD, common bile duct; GB, gallbladder.
3. In Vivo GEM Model of BTC
Owing to
Alb expression in hepatoblasts, which are bi-potential progenitors for hepatocytes and cholangiocytes, liver-specific tumor models have been generated mostly by intercrossing
Alb-Cre mice with conditional GEMs carrying floxed alleles. However, the histology of the resulting tumors varies among HCC, iCCA, and the mixed type, depending on the reconstitution of genetic alterations. For example, in mice with liver-specific homozygous deletion of
Smad4 and
Pten (hereafter
Alb-Cre;
Smad4f/f;
Ptenf/f), all mice developed iCCA and died after 10 months, while
Alb-Cre;
Ptenf/f mice predominantly developed HCC [
100]. In
Alb-Cre;
Ptenf/f;
KrasLSL-G12D/+ mice, only iCCA was observed, while both iCCA and HCC were observed in
Alb-Cre;
Ptenf/+;
KrasLSL-G12D/+ mice [
101]. Considering that many factors can affect the development of iCCA and HCC, it is challenging to generate genuine iCCA or HCC separately by genetic aberrations in a deterministic way. Other mouse models for iCCA were mixed with HCC to varying extents, including
Alb-Cre,
KrasLSL-G12D/+,
Trp53f/f,
Alb-Cre,
KrasLSL-G12D/+,
Ptenf/f,
Alb-Cre;
KrasLSL-G12D/+;
Idh1f/f,
Alb-Cre;
Notch1;
KrasLSL-G12D/+,
Alb-Cre;
KrasLSL-G12D/+;
Fbxw7f/f, and
Alb-Cre;
Cdh1f/f;
Sav1f/f;
Hspd1f/f [
101,
102,
103,
104,
105,
106,
107,
108,
109].
Krt19-Cre is often used to induce genetic alterations in the biliary system, although
Krt19 expression can also be detected in many glandular epithelia and non-keratinized squamous basal cells throughout the body [
110]. In contrast,
Krt19-Cre,
KrasLSL-G12D/+,
Tgfbr2f/f, and
Cdh1f/f mice invariably developed eCCA and
Krt19-Cre,
KrasLSL-G12D/+, and
Tgfbr2f/f mice died from respiratory cancers without eCCA development [
22]. These results suggest that the lack of tissue specificity could impair the relevance of BTC models, especially when other organs become more cancer-prone. There are no known genes specifically expressed in GB. Nonetheless,
Pdx1-Cre, often used to induce pancreatic carcinogenesis, also developed GBC by crossing with
KrasLSL-G12D/+ mice [
111]. Transgenic mice expressing rat
ErbB-2 under the
bovine keratin 5 promoter, which was aimed at developing skin tumors, also developed GBC [
112,
113], underscoring the oncogenic potential of
ErbB-2.
4. Implantation-Based Hybrid Mouse Model of BTC
Although GEM is currently the gold standard in modeling tumorigenesis, it is still impossible to perfectly phenocopy all aspects of sporadic carcinogenesis. For example, currently available Cre mice may not provide sufficient tissue specificity, and the resultant gene recombination is homogenously observed across the target organ during development. In contrast, the tumor originates from a single mutated cell in adults, among many genetically intact normal cells in the organ. To address these issues, novel hybrid models have been developed by combining the in vitro genetic engineering of primary cells and subsequent implantation into host mice in various types of cancers, including BTC, intestine, lung, and pancreas [
19,
58,
59,
88,
114,
115]. This hybrid approach can also integrate GEM as a source of primary cells, thereby accelerating the modeling of BTC without multiple intercrossings to generate many GEMs. The implementation- and organoid-based BTC models are summarized in .
This entry is adapted from the peer-reviewed paper 10.3390/cancers13102292