1. A Role at the Synapse
alpha-Synuclein (aSyn) is a small, intrinsically disordered protein that is mainly localized at the pre-synaptic terminal [
26,
27], but is also present in the neuronal somato-dendritic compartment [
28], in red blood cells [
29], in the gut and other peripheral tissues [
30,
31,
32]. Although aSyn is highly enriched in presynaptic boutons, it displays a delayed distribution in the terminals, suggesting that it is implicated in later stages of synaptic development, rather than playing a central role in synapse modulation [
27]. Importantly, aSyn is differentially expressed in the various neuronal cell types, being more abundant in excitatory synapses across different brain regions and particularly in central catecholaminergic systems [
33]. On the contrary, the protein displays a differential expression profile in inhibitory synapses amongst the different brain areas, with a particular interest of aSyn presence in striatal GABAergic medium spiny neurons (MSNs) [
34,
35].
The first indication regarding the role of aSyn on neural plasticity arose about 25 years ago, when “synelfin” (synuclein, NACP) expression was found up-regulated during bird song learning [
36]. The localization of aSyn in pre-synaptic boutons is mainly attributed to its tight association with synaptic vesicle membranes [
37] and its high affinity for the SNARE complex proteins synaptobrevin-2 (or Vesicle Associated Membrane Protein 2, VAMP2), synapsin III and rab3A [
38,
39,
40]. It has been proposed that aSyn interacts with VAMP2 and promotes SNARE complex assembly [
38], followed then by its disassembly in order to complete the round of membrane fusion (). The crucial role of aSyn assembly with SNARE complex on neuronal survival was further verified by the neuronal dysfunction and impaired survival of triple αβγ-synuclein knockout mice during ageing [
38,
41]. Interestingly, aSyn lentiviral overexpression in primary neurons led to enhanced SNARE complex assembly, further supporting the role of this protein in synaptic activity [
38]. The same group later showed that only multimeric membrane-bound, but not the soluble monomeric aSyn, can promote the SNARE complex assembly [
42]. It has been also recently suggested that aSyn is involved in synaptic vesicle homeostasis at the pre-synaptic terminal via a calcium (Ca
2+)-dependent mechanism [
43].
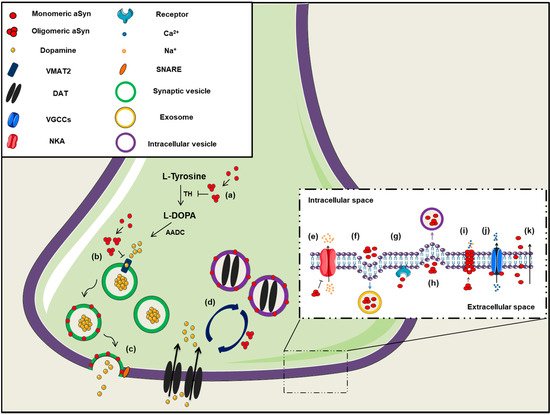
Figure 1. The role of aSyn at the presynaptic terminal. A schematic representation depicting of aSyn physiological and pathological effects at the synapse: (a) aSyn reduces the activity of tyrosine hydroxylase (TH), the enzyme responsible for catalyzing the conversion of L-Tyrosine to L-DOPA, thus impairing dopamine biosynthesis, (b) Increased levels of aSyn inhibit VMAT2, which is responsible for the uptake of monoamines (such as dopamine) into synaptic vesicles, (c) aSyn associates with synaptic vesicle membranes and regulates the SNARE-dependent vesicle fusion and neurotransmitter release, (d) Soluble aSyn interacts with the dopaminergic transporter DAT and decreases its amount on the plasma membrane, thus regulating the dopamine re-uptake from the synapse. However, aSyn aggregates trigger DAT recruitment to the plasma membrane that leads to massive entry of dopamine, (e) aSyn aggregates interact with Na+/K+-ATPase (NKA) preventing the effective pump out of Na+ ions, (f) aSyn is secreted from neuronal cells partly via associating with exosomes, (g) Extracellular aSyn interacts with neuronal receptors (i.e., LAG3) for its internalization in neurons or (h) it is up-taken via endocytosis, (i) PD-linked A30P and A53T mutant aSyn form large membrane pores through which most cations (i.e., Ca2+) can pass non-selectively, (j) Extracellular aSyn activates the voltage-gated Ca2+ channels (VGCCs), resulting in increased Ca2+ influx, (k) Monomeric aSyn enters neuronal cells via passive diffusion or direct penetration of their plasma membrane.
On the contrary, natively unfolded monomeric aSyn at the pre-synaptic terminal is prone to form pathological conformations, thus exerting neurotoxic effects [
44] (). It has been additionally suggested that aSyn is preferably bound to synapsin 1 and VAMP2 when the protein is present in its oligomeric form [
45], highlighting the importance of the conformational state of aSyn for its proper function. There are also findings supporting the implication of aSyn in synaptic transmission, due to its association with the synaptic vesicle pool, modulating the vesicle mobility, the recycling pool homeostasis and endocytosis [
46,
47,
48].
Alpha-synuclein can also function as a molecular chaperone via effective binding to other intracellular proteins. The first indication came with the discovery that aSyn displays structural and functional homology with other molecular chaperones, as the 14-3-3 or small heat shock proteins [
49,
50]. Additional studies revealed that aSyn synergistically acts with the presynaptic cysteine-string protein-alpha (CSPalpha) promoting the assembly of the SNARE complex [
38,
51], further validating its chaperoning properties. Biochemical and structural analysis of aSyn strengthened the current indications for its chaperone-like function via its C-terminal region (residues 61-140) [
52,
53,
54]. However, following studies indicated that the chaperone-binding site of aSyn lies within the non-amyloidal component (NAC) region (residues 61-95), which is prone to aggregation and thus highly susceptible to form fibrils [
55,
56].
2. Association with Membranes and Lipid Trafficking
Intracellular aSyn can be found either natively unfolded in a soluble state or membrane-bound forming an alpha-helical or a beta-sheet secondary structure, depending on the solution conditions [
57,
58,
59]. It has been proposed that there is a bidirectional link between aSyn species formation and membrane remodeling, meaning that not only aSyn structure is affected upon lipid interaction, but also that membrane integrity depends on the presence of different aSyn conformations [
60,
61,
62]. However, there are controversial results regarding the association of aSyn with membrane lipids and its conformational state, with some studies reporting that membrane-bound aSyn gets protected from aggregation, thus leading to neurotoxicity attenuation [
44,
63,
64], whereas others suggest that interaction of aSyn with membranes triggers its self-association and subsequent aggregation [
65,
66,
67]. Importantly, it has been shown that the PD-related aSyn mutations reduce its interaction with membranes, thus further suggesting that aSyn binding on membranes may exert neuroprotective effects [
68,
69,
70,
71,
72].
A plethora of studies argue that aSyn in its soluble state exists as a monomer [
73,
74,
75,
76], whereas others suggest that it occurs physiologically as a tetramer resisting aggregation [
77,
78,
79]. In the presence of lipid membranes, aSyn adopts an alpha helical structure in the N-terminus region that stabilizes the formation of high-order aSyn multimers [
42,
73,
80,
81]. Interestingly, the membrane curvature seems to affect the structure of aSyn, which can adopt either an elongated or a broken alpha-helix conformation, when bound to a large diameter (∼100 nm) or a small, highly curved vesicle, respectively [
82,
83,
84,
85]. It has been also proposed that aSyn has a role in lipid metabolism, since it participates in fatty acids transportation between the cytosol and membranous compartments [
86,
87] and in lipid and membrane biogenesis organizing and stabilizing the lipid bilayer of membranes and vesicles [
88,
89]. On the other hand, disrupted aSyn expression pattern leads to lipid dysregulation, since both the absence and the overexpression of either wild-type (WT) or mutated aSyn gives rise to abnormal lipid metabolism [
90,
91,
92,
93]. Finally, several studies have demonstrated that aSyn regulates membrane homeostasis via inhibition of phospholipases activity, such as phospholipase D [
94,
95,
96,
97]; however, there are controversial results in the literature [
98].
3. Aggregation and Post-Translational Modifications
alpha-Synuclein is composed of three distinct domains: the N-terminal lipid-binding domain, the NAC region and the C-terminal binding domain [
84,
99,
100]. A central role in the fibril formation and subsequent aggregation of aSyn is thought to be mediated through the NAC region of the protein composed of nonpolar side-chains and assembles cross b-structures. Based on that, it has been shown that the deletion of specific residues (74-84) within the core region can abolish aSyn aggregation [
101,
102]. It has been also demonstrated that the endogenous neuronal aSyn and the interaction of aSyn with lipids plays a central role for aSyn recruitment and subsequent seeding of pathology, as it could behave as a core for the formation of insoluble aggregates [
35,
75,
103,
104].
Several mutations in the
Snca gene have been linked to PD pathogenesis, such as the A53T, A30P, E46K, H50Q, G51D, A18P, pA29S and A53E mutations, all located in the N-terminus region [
3,
5,
7,
68,
105,
106,
107]. Most of them are tightly linked to enhanced aSyn aggregation, pathology progression and clinical manifestations in PD. Specifically, A53T and A30P aSyn mutants are natively unfolded, similarly to WT protein. However, at higher concentrations A53T has been shown to accelerate aSyn fibrillization, a critical event in PD pathogenesis [
108,
109,
110]. On the other hand, A30P promotes aSyn oligomerization rather than fibrillization, thus reducing aggregate formation [
109,
111]. The E46K mutation leads to conformational changes of aSyn due to C-terminal to N-terminal contacts in the monomeric protein, resulting in enhanced aSyn accumulation [
111,
112,
113]. Moreover, the PD-linked H50Q point mutation increases aSyn aggregation propensity and toxicity [
114], whereas the G51D mutation has the opposite effects [
115]. However, although G51D mutants seem to oligomerize in a slow rate, they form more toxic fibrils, thus suggesting distinct disease mechanisms for the various aSyn mutations [
116,
117]. Similarly, A53E mutant seems to lead to neuronal toxicity via an aSyn aggregation-independent manner [
118]. Strikingly, the G51D and A53E aSyn mutations have been proposed as potential links between PD and MSA [
106,
119]. However, up-to-date, no hereditable mutations in the coding region of
SNCA gene have been identified in MSA cases [
120]. Apart from point mutations [
117,
121,
122], various post-translational modifications are implicated in aSyn aggregation, the most important of which are phosphorylation, sumoylation, ubiquitination, nitration, N-acetylation, O-GlcNAcylation and truncation.
The phosphorylation of aSyn both at serine and tyrosine residues and particularly at Ser129 is widely considered as an indicator of pathology. However, the effect of Ser129 phosphorylation on aSyn toxicity is still under debate, with the majority of studies suggesting that it accelerates cell toxicity and neurodegeneration [
123,
124,
125,
126,
127]. Contrarily, others have proposed a neuroprotective role of Ser129 phosphorylation since it was reported to drive the conversion of toxic oligomers into less harmful aggregates [
128,
129,
130]. Other mechanisms of phosphorylated Ser129 aSyn-mediated neuroprotection include inhibition of its fibrillation [
131], upregulation of tyrosine hydroxylase (TH) activity [
132] or lowering of the protein’s membrane-binding affinity [
133]. Although the 90% of aSyn in LBs is found phosphorylated at Ser129, a significant amount of phosphorylated Ser129 aSyn is also detected in a soluble, rather than in an aggregated state in PD brains [
134], whereas only a small percentage of aSyn is phosphorylated at Ser129 in the brains of healthy controls [
135,
136,
137]. In addition, aSyn can be phosphorylated at Ser87, Tyr125, Tyr133 and Tyr136 residues [
138,
139] and these are also implicated in either neurotoxic or neuroprotective events [
127,
138,
140,
141]. Nonetheless, in most in vivo models where aSyn is overexpressed (virally, transgenic or PFF-inoculations) the detection of pSer129 positive aSyn signal is invariably linked to neurotoxicity, indicating a rather neurotoxic and not a neuroprotective role.
Nitrated aSyn is also tightly linked to neurodegeneration, as demonstrated by experiments in both cellular and animal models, as well as in patient-derived brains [
142,
143,
144,
145], through its implication in oxidative damage and disease development [
146]. Four tyrosine residues in aSyn sequence, Tyr39 (within the N-terminus), Tyr125, Tyr133 and Tyr 136 (within the C-terminus) can undergo nitration. Nitration at Tyr39 has been shown to result in low binding affinity of aSyn on lipid vesicles due to its loss-of-alpha helical conformation status [
147], whereas nitration at Tyr125 seems to play a crucial role for aSyn dimerization [
148]. Moreover, the linking between two tyrosines is considered as a potential mechanism for aSyn oligomer stabilization and its subsequent aggregation into proteinaceous inclusions [
149]. In addition, the detection of nitrated aSyn in the human blood serum could potentially serve as a clinical biomarker for PD diagnosis [
150].
Another aSyn post-translational modification crucial for its aggregation propensity is ubiquitination, via regulation of the proteasome-dependent protein degradation [
151] and the subcellular localization of the protein [
152]. Ubiquitinated aSyn has been isolated from LBs and sarkosyl-insoluble fractions derived from synucleinopathy brains [
153,
154]. CHIP (C-terminal U-box domain of co-chaperone Hsp70-interacting protein), SIAH (seven in absentia homolog) and Nedd4 (neuronal precursor cell-expressed, developmentally down-regulated gene 4) have been identified among the E3 ubiquitin ligases implicated in aSyn ubiquitination [
155,
156,
157,
158,
159,
160]. Ubiquitin modification has been demonstrated to have differential effects on aSyn accumulation and subsequent aggregation, dependent on the residue being modified. More precisely, ubiquitination at Lys6, Lys12 and Lys21 residues has been shown to moderately inhibit aSyn fibrillation, whereas at Lys10 and Lys23 residues has been reported to promote the formation of aSyn inclusions [
161]. In addition, ubiquitination at Lys32, Lys34, Lys43 and Lys96 inhibits aSyn aggregation [
161].
Sumoylation is a similar process to ubiquitination, since aSyn is conjugated to SUMO (small ubiquitin-like modifier) at lysine residues. SUMO-1 was found in aSyn-positive inclusions of a-synucleinopathy brains or associated with lysosomes of PD animal models [
162,
163,
164]. It has been also suggested that aSyn sumoylation facilitates its aggregation since it inhibits its degradation [
165], whereas other studies proposed a neuroprotective role of aSyn sumoylation, which seems to promote aSyn solubility and thus inhibit its aggregation [
166,
167]. The discrepancy between these data may be attributed to the different lysine residues available for sumoylation being investigated in each study. Another aSyn modification that has been up for debate is its N-terminal acetylation. Although many studies have assigned a neurotoxic role on aSyn N-acetylation, as it has been shown to promote aSyn β-sheet formation and fibrillation [
168,
169,
170], others suggest that either N-acetylated aSyn mediates its physiological binding on synaptic vesicles [
171], or it acts in a protective manner against aSyn aggregation [
172,
173].
O-GlcNAcylation is a biochemical process that involves the attachment of O-linked N-acetylglucosamine to Ser and Thr residues of various proteins, amongst which is aSyn. Murine and human aSyn have been shown to be O-GlcNAcyled in many threonine residues including Thr33, Thr34, Thr54, Thr59, Thr64, Thr72, Thr75, Thr81 and Thr87 [
174,
175,
176,
177,
178] and this post-translational modification has repetitively been linked to reduced aSyn aggregation and attenuation of PD-related toxicity [
179,
180,
181,
182]. Finally, aSyn truncation has gained scientific attention, given that C-terminally truncated aSyn has been identified in the inclusions present in PD brains [
183,
184,
185]. Many studies have considered that aSyn truncations have neurotoxic effects due to increased accumulation of misfolded aSyn [
186,
187,
188,
189,
190,
191,
192,
193,
194].
4. Channel Formation/Channel Interactions
As mentioned above, membrane-bound aSyn adopts an alpha-helical conformation, which facilitates its oligomerization and subsequent aggregation. It has been suggested that aSyn oligomers can form transmembrane channels and pore-like structures that have been linked to pathological events during PD development () [
195,
196,
197]. As a result, vesicles or low-molecular mass molecules may penetrate the cell membrane and in combination with altered cellular ionic homeostasis could potentially lead to cell toxicity and neuronal degeneration [
198,
199]. Another mechanism for the increased membrane permeability involves the incorporation of aSyn oligomers between the membrane phospholipids, thus leading to the bilayer thinning which thereafter allows the diffusion of small molecules [
200].
A wide range of studies has demonstrated that the ion channels formed by oligomeric aSyn dysregulate cellular ion concentrations and may represent a critical event in the pathogenesis of a-synucleinopathies [
198]. Some PD-linked aSyn mutations, such as E46K and A53T, have been shown to be implicated in the channel formation, whereas other aSyn mutants (i.e., A30P) have displayed low membrane affinity [
197,
201]. However, other groups have shown that A30P and A53T aSyn mutations are responsible for the formation of large membrane pores through which most cations can pass non-selectively [
202]. It has been reported that the formation of such cation-permeable pores could lead either to ion conductivity or to increased Ca
2+ influx and subsequent cell death [
202,
203,
204,
205]. Upon aSyn cation channel opening, other channels, such as the ATP-dependent potassium channels K (ATP), have been reported to be activated in hippocampal neurons and this could probably diminish the aSyn-dependent neuronal excitability [
205].
Binding of aSyn to the plasma membrane results in the formation of aggregates and this aggregation leads to the redistribution of the α3 subunit of Na
+/K
+-ATPase. As a result, Na
+/K
+-ATPase is no longer able to effectively pump out Na
+ from neurons, thus leading to an intracellular Na
+ accumulation [
206]. Furthermore, extracellular aSyn was reported to activate the voltage-gated Ca
2+ channel Cav2.2 in rat neurons, due to disorganization of lipid rafts in the plasma membrane, resulting in enhanced dopamine release and increased Ca
2+ influx [
207]. Both events may explain the synaptic dysfunction and neuronal vulnerability in PD. L-type Ca
2+ channels are also implicated in PD development, as administration of L-type Ca
2+ channel blockers (i.e., isradipine, nimodipine) in animal models and PD patients, reduced death risk and ameliorated disease manifestations [
208,
209,
210,
211]. Finally, aSyn oligomers can inhibit α4β2 nicotinic acetylcholine receptors of dopaminergic neurons, thus leading to cholinergic signaling deficits [
212]. In summary, aSyn seems to regulate neuronal toxicity and survival via the formation of channels or pores in the plasma membrane or via its interaction with other channels or receptors crucial for the proper neuronal activity ().
6. Interaction with Mitochondria and ER
alpha-Synuclein displays a remarkable conformational flexibility upon macromolecular interactions and can associate with mitochondrial membranes, thus altering mitochondrial function [
230,
231,
232] (). There are reports suggesting that aSyn is a physiological regulator of mitochondrial activity [
233,
234,
235], whereas others support the opposite [
236,
237,
238]. Such discrepancies could be attributed to the different synuclein models utilized in each study, taking into account that brain homeostasis is a complex process and in vivo studies recapitulate better the interplay between the various brain components, compared to the isolated in vitro cellular setup. A bidirectional interaction between aSyn aggregation and mitochondrial dysfunction has been implicated in PD pathogenesis. In particular, increased levels of aSyn can lead to mitochondrial dysfunction [
239,
240,
241,
242,
243,
244], whereas, conversely, impairment of mitochondrial activity may accelerate aSyn pathology [
245,
246,
247,
248]; however, the precise underlying mechanisms remain to be elucidated. Both WT and mutant aSyn have been shown to interact with mitochondrial elements, altering both mitochondria morphology and function. Specifically, soluble pre-fibrillar aSyn oligomers seem to be responsible for complex I dysfunction, loss of membrane potential, disrupted Ca
2+ homeostasis, enhanced cytochrome c release and ROS production, thus leading to neuronal demise [
240,
249,
250,
251,
252].
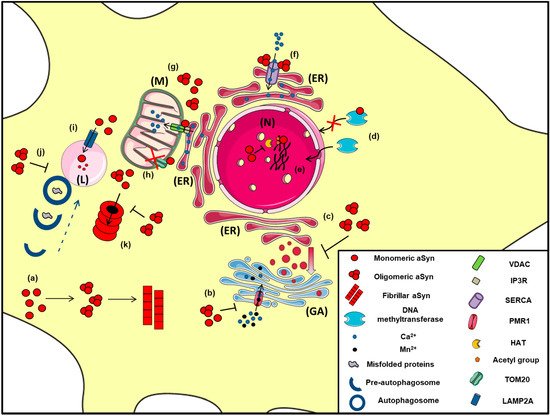
Figure 2. The proposed intracellular effects of various aSyn conformations in neurons. A schematic representation of the aberrant interactions between the various aSyn species with intracellular organelles: (a) In the cytoplasm of neurons, aSyn monomers form oligomers that can eventually become fibrils, (b) Both unfolded and aggregated aSyn impair the function of PMR1, a Ca2+-transporting ATPase pump that regulates Ca2+ and Mn+2 levels in the Golgi apparatus (GA), resulting in elevated cytosolic Ca2+ levels, (c) Both WT and mutant A53T aSyn disrupt the vesicular transport from Endoplasmic Reticulum (ER) to Golgi (GA), (d) WT aSyn inhibits the transportation of methyltransferases from the cytoplasm to the nucleus (N), thus altering DNA methylation of the SNCA gene, (e) Inside the nucleus (N), aSyn inhibits histone acetylation via its direct binding to histones or by preventing the action of histone acetyltransferase (HAT) enzymes, thus interfering in the process of gene transcription, (f) In the ER, aSyn aggregates activate the Ca2+-ATPase SERCA, resulting in dysregulated Ca2+ homeostasis, (g) Both monomeric and oligomeric aSyn interact with Voltage-dependent anion channel 1 (VDAC1) and inositol triphosphate receptors (IP3Rs), the protein components involved in mitochondrial-associated ER membrane (MAM) and regulates the transmission of Ca2+ signals from the ER to mitochondria (M), (h) aSyn binds to TOM20, a mitochondrial import receptor subunit and inhibits normal protein import, (i) Normally, monomeric or dimeric forms of aSyn are degraded in the lysosome (L) via Chaperone Mediated Autophagy (CMA), following their interaction with LAMP2A. However, under pathological conditions, impairment of CMA has been proposed to lead to aSyn accumulation and subsequent cell toxicity, (j) Oligomeric aSyn and various misfolded proteins are cleared via macroautophagy, following the fusion of autophagosomes with the lysosome. Pathological aSyn has been shown to inhibit autophagosome maturation or their fusion with lysosomes, thus impairing autophagic flux, (k) Monomeric and oligomeric aSyn are degraded via the proteasome; however, under pathological conditions, increased levels of aSyn or even soluble aSyn oligomers may inhibit proteasomal function, leading to aSyn accumulation and the formation of insoluble aggregates.
Experiments in various animal models of a-synucleinopathy have revealed mitochondrial abnormalities, DNA damage and neuronal degeneration in PD-affected brain regions [
244,
253,
254]. Moreover, in vitro and in vivo experiments have shown that aSyn inhibits mitochondrial fusion and triggers mitochondrial fragmentation [
231,
255]. Di Maio and colleagues have proposed that certain post-translationally modified aSyn conformations (soluble oligomers, dopamine-modified and S129E phosphorylation mimic) lead to impaired mitochondrial function via binding to TOM20 (translocase of the outer membrane receptor) and inhibiting mitochondrial protein import [
239].
Nonetheless, there is evidence suggesting an impairment of mitochondrial function upstream of aSyn pathology. Experiments using the pesticides rotenone and paraquat have shown that dysregulation of mitochondrial function leads to nigrostriatal dopaminergic loss and formation of LB-like inclusions, positively stained with anti-aSyn antibodies and thioflavine S, thus resembling PD features [
246,
247,
256,
257,
258]. Similarly, incubation of WT aSyn-overexpressing COS-7 cells with mitochondrial inhibitors resulted in the disappearance of the aSyn aggregates formed upon rotenone or oligomycin treatment [
259]. A plethora of studies that utilize the mitochondrial neurotoxin MPTP to induce PD-like pathology in animals, further suggest that mitochondria impairment is a key player in disease development [
245,
248,
260,
261,
262,
263,
264]. Genetic studies further support the hypothesis of aSyn accumulation as a secondary event following mitochondrial malfunction. Specifically, mutations in ATP13A2 (ATPase cation transporting 13A2), encoding for the lysosomal type 5 P-type ATPase, were shown to result in dysregulation in mitochondrial depolarization and ATP metabolism leading to mitochondrial fragmentation and subsequent cell death [
265,
266].
Apart from its implication in mitochondrial failure, aSyn has been also reported to play a biological role in the association of mitochondria with the endoplasmic reticulum (ER) Ca
2+ homeostasis. It has been demonstrated that aSyn favors the Ca
2+ transfer from ER to mitochondria, as a result of the communication the two organelles, probably due to the fact that aSyn can act as a “bridge” via its C terminus [
267]. Later studies further supported the physiological localization of aSyn in mitochondria-associated ER membranes (MAM), stabilizing their interaction, which was perturbed upon aSyn aggregation and its subsequent redistribution [
268,
269]. Interestingly, the familial PD-linked A53T and A30P aSyn point mutations resulted in their weakened interaction with MAM, which affected MAM function and mitochondrial integrity [
269].
The association of aSyn with mitochondria was further corroborated by findings indicating interactions between both monomeric and oligomeric aSyn with the Ca
2+ transporting voltage-dependent anion channel 1 (VDAC1) [
270,
271,
272,
273]. Importantly, VDAC1 has been detected on the MAM of ER mediating the communication between the two organelles, regulating Ca
2+ homeostasis [
274,
275,
276]. Moreover, VDAC levels have been found decreased in nigral neurons of PD brains, where pathological aSyn inclusions had been formed [
277]. Additionally, VDAC has been proposed to be a component of the mitochondrial permeability transition pore, the opening of which has been shown to be affected by aSyn overexpression and oligomerization [
230,
278]. In vivo experiments on transgenic mice overexpressing the human A53T aSyn further supported the role of permeability transition pore activity modulation on the mitochondrial dysfunction during PD pathogenesis [
279].
7. Unfolded Protein Response, Regulation of ER/Golgi Trafficking and Ca2+ Homeostasis
The ER is a continuous membrane system mainly responsible for the production and processing of lipids and proteins, as well as Ca
2+ homeostasis. In case of impaired protein folding (ER stress), cells activate a group of signal transduction pathways, known as the unfolded protein response (UPR). It has been previously shown that aSyn overexpression in PD patients leads to UPR and contributes to the molecular pathogenesis of the disease [
280]. The ER chaperone glucose regulated protein 78 (GRP78/BIP) has a crucial role on ER stress regulation due to its ability to control the activation of transmembrane ER stress sensors (IRE1, PERK and ATF6) [
281]. Disassociation of GRP78 from IRE1 and PERK results in stress signaling, finally leading to altered ER homeostasis [
282]. aSyn associates with GRP78/BIP under physiological or pathological conditions, thus inducing UPR and leading to dopaminergic cell death [
45,
283]. Strikingly, Ser129 phosphorylated and aggregated aSyn was found in ER microsomes of A53T transgenic mice and more importantly, administration of the UPR inhibitor salubrinal, effectively attenuated disease manifestations in this PD-mouse model [
284,
285]. It is worth mentioning that GRP78/BiP levels were found elevated in DLB and PD brains in an aSyn burden-dependent manner [
286]. In addition, the protein levels of various ER chaperones were found elevated in a-synucleinopathy models, co-localized with aSyn positive inclusions, suggesting that aggregated aSyn could potentially be implicated in UPR regulation in disease progression [
284,
287,
288,
289,
290,
291,
292,
293].
Proteins synthesized in the ER, are packaged into vesicles and directed to Golgi apparatus for subsequent modifications. One of the first pathological roles attributed to aSyn is the blockade of the vesicular transport from ER to Golgi by antagonizing ER/Golgi SNAREs [
294,
295,
296]. Towards the same direction, aSyn can also disrupt the intra-Golgi and post-Golgi secretory trafficking, via an abnormal interaction with several Rab-family proteins of the intracellular endocytic pathway [
294,
296,
297,
298,
299]. Additionally, aSyn can also impair the ionic transport and membrane trafficking, resulting in Golgi fragmentation and subsequent cytotoxicity [
300,
301,
302].
Another significant role of aSyn on ER and Golgi function is the regulation of Ca
2+ homeostasis via its binding on specific channels or pumps localized in these organelles (). Specifically, proximity ligation assay experiments demonstrated that soluble and insoluble aSyn aggregates, but not monomers, interact with the ER Ca
2+-ATPase SERCA, resulting in decreased cytosolic Ca
2+ that disrupts the physiological cell function and leads to neuronal cell death [
303]. Moreover, administration of the SERCA inhibitor cyclopiazonic acid restored cytosolic Ca
2+ levels and protected neurons against the aggregated aSyn-dependent cell death [
303]. In support to these results, aggregated aSyn bound on SERCA pump was detected in LBs and GCIs of PD and MSA brains, respectively [
303]. Furthermore, PMR1, a Ca
2+-transporting ATPase 1 pump regulating the levels of Ca
2+ and Mn
+2 ions in the Golgi [
304], has been proposed to be a mediator of aSyn-dependent cytotoxicity. Specifically, in various PD models (yeast, flies and nematodes), PMR1 pump has been linked to aSyn pathology via a Ca
2+-dependent mechanism, where aSyn accumulation elevated cytosolic Ca
2+ levels and increased cell death. Interestingly, upon PMR1 deletion, the disease-associated characteristics were abolished, further suggesting the relevance of this pump to aSyn pathology [
305,
306].
8. a-Synuclein in the Nucleus
The name aSyn was given to the protein due to its localization in the nucleus and presynaptic nerve terminals [
37]. Nuclear aSyn was detected in neurons of various brain regions of rodents and was reported to interact with histones, underlying PD pathology [
307,
308,
309], even though a single study declares that the nuclear staining of aSyn is attributed to the non-specific signal of some antibodies that probably recognize unknown antigens in neuronal nuclei [
310]. It has been proposed that aSyn is responsible for epigenetic dysregulation via inhibition of histone acetylation or reduced DNA methylation, thus favoring neuronal degeneration, whereas others suggest that nuclear aSyn regulates cell cycle rate exhibiting cell toxicity [
311,
312,
313]. Importantly, histone deacetylase (HDAC) inhibitors attenuated aSyn toxicity and provided neuroprotection in both cell culture and transgenic Drosophila models [
311,
314].
Experiments in SH-SY5Y cells revealed that nuclear translocation of aSyn is regulated by calreticulin and Ca
2+, following treatment with retinoic acid and modulates the expression of PD-linked genes such as ATP13A2 and PINK1 (PTEN-induced kinase1) [
315]. Interestingly, phosphorylated aSyn at Ser129 was found accumulated in the nucleus of HEK293E-aSyn overexpressing cells and in various brain regions of transgenic (Thy1)-[A30P] aSyn mice [
316]. Further experiments in H4 cells expressing various aSyn proteins verified that nuclear localization of aSyn depends on its phosphorylation at Ser129 [
317]. The same group supported a role of DNA-binding and gene expression regulation for aSyn providing an insight into the role of modified aSyn in the nucleus [
317]. Furthermore, other post-translational modifications of aSyn, such as sumoylation, seem to be responsible for the translocation of aSyn from the cytoplasm to the nucleus [
318]. Although the majority of studies support a neurotoxic role for aSyn nuclear localization, some groups proposed that aSyn in the nucleus displays a protective role against DNA damage, replication stress or impaired nucleo-cytoplasmic transport [
319,
320,
321]. However, the numerous in vitro and in vivo studies demonstrating a neurotoxic role of nuclear aSyn, in contradiction to the limited number of studies supporting a protective role originated mostly from cell lines or yeast, favors the pathological potential of nuclear aSyn.
9. Alpha-Synuclein and Protein Degradation Pathways: An Intricate Interplay
A great wealth of data focuses on the complicated relationship between aSyn clearance and protein degradation pathways (). Both the ubiquitin-proteasome system (UPS) and the autophagy lysosome pathway (ALP) are responsible for aSyn degradation in a manner that depends on cell type, tissue and aSyn conformation state [
322,
323,
324]. Specifically, there are studies demonstrating that aSyn can be degraded by the 26S/20S proteasome via ubiquitin-dependent [
325,
326] and ubiquitin-independent manner [
327,
328]. Studies in PC12, HEK293 and primary mesencephalic cells suggested that pharmacological inhibition of the proteasome does not lead to aSyn accumulation [
324,
329,
330]; however, others have shown that soluble aSyn oligomers, but not monomers, are partially cleared via the 26S proteasome [
331]. Importantly, it has been proposed that the UPS is responsible for aSyn removal under normal conditions, while in pathological cases the ALP is recruited to clear the increased aSyn burden [
332].
Chaperone-mediated autophagy (CMA) is also responsible for the degradation of monomeric or dimeric forms of the protein via the lysosome-associated membrane protein type 2A (LAMP2A), whereas oligomeric aSyn is cleared mainly via macroautophagy [
324,
333,
334]. Lee and colleagues also suggested that the lysosome is responsible for the removal of oligomeric but not fibrillar aSyn and that lysosomal failure results in aSyn accumulation and aggregation and subsequent cell death [
335]. Moreover, initial in vivo evidence suggested that increased aSyn protein levels evoked by paraquat treatment were preferably degraded via CMA in dopaminergic neurons, where the levels of LAMP2A and the lysosomal heat shock cognate protein of 70 kDa (HSC70), both essential CMA-components, were found elevated [
336]. We have also shown that boosting CMA function via LAMP2A overexpression in cell lines and primary neuronal cultures and in the rat dopaminergic system mitigated aSyn protein levels and related toxicity [
337]. Similar neuroprotective effects were obtained upon LAMP2A overexpression in the Drosophila brain [
338]. On the contrary, we have also shown that LAMP2A silencing led to endogenous aSyn accumulation in vitro [
324] and in vivo [
339] and in extensive neurodegeneration of the rat nigrostriatal axis [
339]. Decreased levels of LAMP2A and HSC70 were reported in the human substantia nigra and amygdala of PD brains [
340], whereas, in a subsequent study, LAMP2A was found to be selectively reduced in association with increased aSyn levels, even in the early stages of PD, thus suggesting a potential dysregulation of CMA-mediated protein degradation prior to substantial aSyn aggregation in PD [
341].
However, a bidirectional link between aSyn accumulation and the protein degradation machineries exists and extensive studies have been conducted to elucidate not only the manner of aberrant aSyn degradation in a-synucleinopathies, but also the impact of various aSyn conformations on UPS and ALP function. It has been proposed that overexpression of A30P and A53T mutants, contrarily to WT aSyn, leads to cell death due to proteasomal inhibition [
342]. Indeed, overexpression of mutant A53T aSyn resulted in UPS failure by inhibiting the activity of the 20S/26S proteasome, finally leading to aSyn pathological accumulation [
343]. Other groups have failed to detect alterations in the proteasomal function of PC12 cells or transgenic mice, following overexpression of WT or mutant (A30P, A53T) aSyn [
344]. Moreover, later studies demonstrated that transient overexpression of WT or mutant aSyn, followed by addition of recombinant aSyn oligomers and fibrils in an osteosarcoma cell line, did not result in any disturbance of the proteasomal function [
345]. Importantly, studies in human post-mortem PD brains also suggested impaired proteasomal function in the substantia nigra [
346,
347,
348], further supporting a role of UPS malfunction in PD pathogenesis. In addition, total rates of protein degradation declines with aging, thus contributing to the pathogenesis of age-related diseases [
349]. Even though human post-mortem studies provide valuable information in regards to etiology and/or disease pathogenesis, the data obtained should be treated with caution, given into account the overall decline in the function of multiple systems with aging. For a-synucleinopathies, we believe that the use of tissue from affected and non-affected (in regards to aSyn pathology and neuronal death) brain areas may provide useful information regarding early or late events leading to neurodegeneration.
Increased aSyn protein burden is reported to impair macroautophagy function as well, via its interaction with Rab1a, an event that subsequently results in the autophagosome-formation-related protein Atg9 mislocalization [
350]. Similar results were obtained from cells expressing the PD-linked mutation of the retromer protein VPS35, which is involved in autophagy and is implicated in PD pathogenesis [
351]. The three most well studied PD-linked aSyn mutations, E46K, A30P and A53T, have been shown to promote ALP dysfunction, via either impairing autophagosome formation or inhibiting the selective removal of damaged mitochondria through mitophagy [
352,
353,
354]. It has been previously reported that dopamine-modified aSyn inhibits CMA and this could probably shed light into the selective vulnerability of dopaminergic neurons in PD [
355]. Further experiments in human iPSC-derived midbrain dopaminergic neurons revealed that disrupted hydrolase trafficking, due to aSyn overexpression, reduces lysosomal function [
356]. Similarly, multiple studies suggest that there is a strong relationship between decreased β-glucocerebrosidase (GCase) activity and aSyn accumulation. In particular, heterozygote mutations in
GBA1 gene encoding for β-glucocerebrosidase represent a major risk factor for PD development with a-synucleinopathy [
357,
358,
359,
360,
361,
362].
10. Alpha-Synuclein in the Extracellular Space
The first indication that aSyn can be secreted arose from the detection of the protein in human CSF and plasma of PD patients, indicating that aSyn can be released into the extracellular space [
363,
364] and can exert various deleterious effects on neighboring cells. Further studies supported that aSyn can be secreted from neuronal cells, either via vesicles or exosomes [
365,
366,
367]. Extracellular aSyn has been the subject of intensive research in recent years, mainly due to its propensity to spread from neuron to neuron or other glial cells, as discussed in the following sections.
The major hypothesis regarding the onset and spread of aSyn pathology in a-synucleinopathies relies in the protein’s nucleation propensity that leads to the formation of aberrant aSyn species, which then spread to neighboring cells and tissues via various mechanisms. Furthermore, aSyn has been proposed to act as a “prion-like” protein since it was demonstrated that pathogenic aSyn could transfer from diseased neurons of a PD patient to the healthy transplanted ones, fourteen years after the surgical intervention [
368]. Similar results were obtained by other groups in both humans and rats [
369,
370,
371,
372]. Experiments of PD and DLB patient-derived brain extracts delivered into the brain of mice and non-human primates further validated the transfer of pathological aSyn and the formation of aSyn aggregates within the recipient neurons [
373,
374]. Moreover, when Pre-Formed Fibrils (PFFs) were used as seeds in both in vitro and in vivo experiments, the endogenous neuronal aSyn was recruited into the formation of highly insoluble aggregates [
104,
375,
376,
377,
378,
379].
Various mechanisms have been proposed for aSyn spread throughout the nervous system, following its release from neurons where the protein is normally expressed. Candidate mechanisms include aSyn secretion via vesicles, exosomes or even naked protein [
364,
365,
366,
380,
381,
382,
383] and its uptake from the cells via conventional endocytosis [
384,
385], passive diffusion [
386], tunneling nanotubes [
387], membrane penetration [
195,
388,
389] or receptor-mediated internalization [
206,
390,
391]. Once taken-up by recipient cells, the exogenous aSyn has been shown to trigger the endogenous aSyn accumulation via an unknown mechanism [
392,
393,
394,
395]. However, according to the prevailing hypothesis, upon the cell-internalization of aberrant aSyn conformations (oligomers or fibrils), these serve as a template for the recruitment of the endogenous monomeric aSyn into the formation of insoluble aggregates [
373,
375,
376,
377,
396,
397,
398]. The prevalently unfolded or alpha-helical aSyn is triggered to self-assemble generating fibrils that subsequently deposit as Lewy bodies [
399,
400,
401].
Neuron-to-neuron aSyn transmission occurs following both anterograde and retrograde axonal transport or trans-synaptic pathways [
402,
403,
404]. Several groups have proposed that dysregulation of axonal transport is implicated in aSyn accumulation at the cell body; however, it is not clear whether PD-linked aSyn mutations play a key role in the process per se [
403,
405,
406,
407]. Notably, aSyn in its oligomeric form has been shown to interfere with microtubules and kinesin motors, thus disrupting the anterograde transport and similar results were obtained in an aSyn overexpressing mouse model for PD, as well as in patients diagnosed with the disease [
408,
409,
410]. Additionally, it has been suggested that the variety in a-synucleinopathy phenotypes is attributed to the formation of different aSyn “strains” that display “aggressive” characteristics [
17,
18,
411]. As a consequence of their disparate structures, these “strains” have discrete biochemical responses along the different brain regions and cell types, thus explaining the various disease manifestations of a-synucleinopathies [
19,
20,
125,
412,
413,
414].