Isoflavones (including daidzein, the glycoside forms of daidzein, and glycitein, the methoxylated form of daidzein) are bioactive compounds that are present in significant quantities in legumes, soybeans, green beans, and mung beans.
- daidzein
- equol
- reproduction
- ovarian folliculogenesis
1. Provenance and Properties of Daidzein and Its Metabolite Equol
Isoflavones (including daidzein, the glycoside forms of daidzein, and glycitein, the methoxylated form of daidzein) are bioactive compounds that are present in significant quantities in legumes, soybeans, green beans, and mung beans. In grains (raw materials), they are mostly present as simple and complex O-glycosides, β-glucuronides and sulfate esters, and highly polar, water-soluble compounds. They are hardly absorbed by the intestinal epithelium and have insignificant biological activities. Beans are included in various food products owing to their digestibility, taste, bioavailability of nutrients, and higher amounts of bioactives. Processing steps include steaming, cooking, roasting, and microbial fermentation, which destroys protease inhibitors and cleaves the glycoside bond to yield absorbable and bioactive aglycone in the processed products, such as miso, natto, soy milk, tofu, and increases the product’s shelf life. Results are reliable (including the conversion of glucoside daidzein to aglicon daidzein, which hydrolyzes conjugated isoflavones with gut bacteria) [2,3,4,6]. Clinical studies show the differences between the biological activities of aglycone and conjugated forms of daidzein [2]. Processed soy food products were an integral part of diets in many Asia-Pacific countries for centuries, but in the last two decades, soy products were successfully integrated into Western diets because of their health benefits [3].
Daidzein (7-hydroxy-3-(4-hydroxyphenyl)-4H-chromen-4-one) (PubChem ID 5281708, molecular formula C15H10O4; chemical structure indicated in Figure 1a, https://pubchem.ncbi.nlm.nih.gov/ (accessed on 16 April 2021)) is an isoflavonoid derived from L-phenylalanine. Daidzein is an aglycone form of daidzein. Its solubility in water is relatively low; therefore, for medical purposes, it is dissolved in alcohol or an alcohol/water mixture.
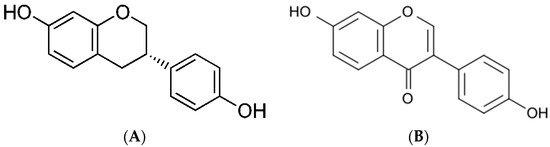
The primary biological and health effect of daidzein is because of its metabolite, equol ((3S)-3-(4-hydroxyphenyl)-7-chromanol) (PubChem ID, 6950272 molecular formula C15H14O3; chemical structure indicated in Figure 1b, https://pubchem.ncbi.nlm.nih.gov/ (accessed on 16 April 2021)). Equol is an isoflavandiol nonsteroidal estrogen and a secondary metabolite of daidzein that is the result of daidzein hydrolysis by gut bacteria. It is insoluble in water and acid-labile [5,13,14]. The conversion of daidzein into equol takes place in the intestine via the action of reductase enzymes belonging to incompletely characterized members of the gut microbiota. While all animal species analyzed so far produce equol, only between one-third and one-half of humans (depending on the community) can do so; ostensibly, those that harbor equol-producing microbes. Only 20%–30% of Westerners produce S-equol compared to 50%–70% of Asians; these subjects might be the only ones who can fully benefit from soy or isoflavone consumption [4,5,13]. Besides microflora, raw plant polyphenols can be metabolized by hepatocytes, intestines, and some other cells, including breast tumor cells. After daidzein enters the circulation via intestinal absorption, it may be transported to the liver, where it is conjugated by liver enzymes to form sulfated, glycosylated, and glucuronidated isoflavones, which are more water-soluble [2,3,6]. Additionally, inflammatory cells produce chemical oxidants that can react with polyphenols and inactivate them [6].
Although isoflavones are nonsteroidal, they have similarities in their chemical structure to that of mammalian estrogens, show estrogenic activities in biological assays, and induce estrogen-like effects in mammalian systems (see below).
Despite the low oral bioavailability, most polyphenols have significant biological effects that highlight the low bioavailability/high bioactivity paradox. This phenomenon can be explained by the high affinity and sensitivity of estrogen receptors, even to a low amount of phytoestrogen [8].
2. Mechanisms of Action of Daidzein and Its Metabolite Equol
3. Physiological and Therapeutic Actions of Daidzein and Its Metabolite Equol
4. Effects of Daidzein and Its Metabolite Equol on Female Reproductive Processes
5. Hormonal Mechanisms of Daidzein and Equol Effects on Female Reproductive Processes
Female reproductive functions, like other functions, are controlled by a hierarchical system of exogenous and endogenous regulators—regulatory molecules produced in the hypothalamus and other areas of the central nervous system (CNS), pituitary hormones (mainly gonadotropins), hormones produced by the ovaries and other peripheral organs, receptors to hormones, and intracellular mediators of their action—and enzymes including protein kinases, transcription factors regulating cell proliferation, apoptosis, viability, and differentiation [34]. The influence of daidzein on some (but not all) of these regulatory molecules is documented.
In the available literature, we failed to find any evidence for daidzein action on the CNS. Furthermore, histomorphological studies did not reveal daidzein influence on morphophysiological indexes of activity in pituitary gonadotropes and lactotropes [26], indicating that daidzein does not affect gonadotropin- and prolactin-producing pituitary cells.
Feeding with daidzein increased the number of follicle stimulating hormone (FSH) and luteinizing hormone (LH) receptors in chicken ovarian follicles [25]. Furthermore, daidzein promoted the stimulatory action of LH on mouse ovarian-cancer development [27]. These observations indicate that daidzein can promote the reception and effect of gonadotropins on ovarian functions. The ability of daidzein to promote the generation of gonadotropin receptors, which was associated with an increase in ovarian folliculogenesis and the egg-laying rate [25] mentioned above, suggests that daidzein promotes ovarian-cell function through the upregulation of gonadotropin receptors. The exact mechanism remains unknown; however, estrogen via estrogen receptors can boost the production of gonadotropin receptors in the ovary [34]. Daidzein can promote the generation of estrogen receptors (see below) [35,36], and mimic gonadotropin action on ovarian cell proliferation [24]. This indicates that daidzein can promote the production of gonadotropin reception through the activation of estrogen receptors. Nevertheless, this hypothesis requires experimental validation.
The action of daidzein on peripheral hormones, including hormones of ovarian origin regulating reproductive functions, is well-documented, although substantial differences are evident in the character of daidzein action, even in similar experiments.
In the in vivo experiments of Medigović et al. [26], injections of diosgenin increased progesterone and estradiol, and decreased testosterone levels in rat serum. On the other hand, in the in vivo experiments of Lamartiniere et al. [18], feeding rats with daidzein reduced the concentration of circulating progesterone but not estrogen.
Some in vitro experiments demonstrated a direct inhibitory action of daidzein and its metabolite equol on steroid hormones‘ release by ovarian cells. Equol inhibited the expression of steroidogenic enzymes, and the synthesis and release of progesterone, testosterone, androstenedione, and estradiol by cultured murine ovarian follicles [23]. Similarly, there was an inhibitory influence with daidzein on progesterone, but not the estradiol release of cultured porcine ovarian granulosa cells, as observed in the in vitro experiments of Nynca et al. [34,35]. Similar experiments by Sirotkin et al. [31] failed to detect daidzein action on progesterone and estradiol release by cultured porcine granulosa cells, but in these experiments, daidzein promoted testosterone output. Other experiments on porcine granulosa cells showed the upregulation of progesterone, but not testosterone and estradiol production [30]. Mlynarczuk et al. [29] did not observe any influence of daidzein on progesterone and estradiol release by cultured bovine granulosa and luteal cells. In their experiments, daidzein was a potent stimulator of enzymes responsible for oxytocin synthesis and release.
Epidemiological studies [37] showed an association between decreased serum daidzein levels and decreased female serum adiponectin levels, and increased serum insulin levels, which could be associated with an elevated risk of ovarian cancer.
The similarity of daidzein and estrogen effects on rat ovarian folliculogenesis [22] and steroidogenesis [26] suggests there is a similar mechanism of action with these molecules on adult ovarian functions via estrogen receptors. This hypothesis was confirmed by the ability of both daidzein and estradiol to bind to estrogen receptors in ovarian and nonovarian tissue [2,3], and to upregulate the expression of estrogen receptors alpha and beta in porcine ovarian cells [35,36]. Moreover, the blockage of estrogen receptors prevented daidzein action on the proliferation of chicken ovarian germ cells [24]. On the contrary, there are reports [27,38] that daidzein can antagonize estrogen action on ovarian-cancer cells. Reports indicate that daidzein affects ovarian-cancer development via both the up- and the downregulation of estrogen receptors. The current literature does not contain direct evidence concerning daidzein action on and via receptors to other peripheral hormones regulating female reproductive processes, but such action cannot be excluded.
Taken together, available data demonstrate the ability of daidzein and equol to regulate the release of steroid and nonsteroid hormones (progestogen, androgens, estrogens, oxytocin, adipokines), and to generate the production of gonadotropin and estrogen receptors. These signaling molecules could be constituents of the integrated endocrine pituitary–gonadal axis. For example, the production of progestogens, androgens, estrogens, and oxytocin is regulated by gonadotropins, while there is also the mutual stimulation of steroid hormones and oxytocin. The gonadotropin–ovarian hormone axis and receptors to these hormones are known regulators of basic ovarian functions and fecundity [34].
Therefore, available data demonstrate that daidzein can affect female reproductive processes via the system of extracellular self-stimulating hormones of pituitary, ovarian, and nonovarian origins and of their receptors. On the other hand, the involvement of only gonadotropin and estrogen receptors in mediating daidzein action is not yet indicated.
This entry is adapted from the peer-reviewed paper 10.3390/ph14040373