Presynaptic Ca2+ entry occurs through voltage-gated Ca2+ (CaV) channels which are activated by membrane depolarization. Depolarization accompanies neuronal firing and elevation of Ca2+ triggers neurotransmitter release from synaptic vesicles. For synchronization of efficient neurotransmitter release, synaptic vesicles are targeted by presynaptic Ca2+ channels forming a large signaling complex in the active zone. The presynaptic CaV2 channel gene family (comprising CaV2.1, CaV2.2, and CaV2.3 isoforms) encode the pore-forming α1 subunit. The cytoplasmic regions are responsible for channel modulation by interacting with regulatory proteins.
- Ca2+ channels
- synaptic transmission
- G-proteins
- synaptic proteins
- Ca2+ binding proteins
1. Introduction
Presynaptic Ca2+ entry into the active zone (AZ) occurs through voltage-gated Ca2+ (CaV) channels which are activated membrane depolarization and triggers synchronous neurotransmitter release from synaptic vesicles (SVs). Multiple mechanisms regulate the function of presynaptic Ca2+ channels [1,2,3,4]. The channel activity for opening, closing, or inactivation in response to membrane depolarization changes every few milliseconds during and after neuronal firing, resulting in control of synaptic strength [3,4]. Following a brief overview of Ca2+ channel structure/function, this article reviews the molecular and cellular mechanisms that modulate the activity of presynaptic Ca2+ channels in the regulation of neurotransmitter release and in the induction of short-term synaptic plasticity. To understand the physiological role of Ca2+ channel modulation in the regulation of synaptic transmission, a model synapse formed between sympathetic, superior cervical ganglion (SCG) neurons in culture was employed for functional study of channel interaction with G proteins, SNARE proteins, and Ca2+-binding proteins which sense residual Ca2+ in the AZ after the arrival of an action potential (AP).
2. Presynaptic Ca2+ Channels
Ca2+ currents have diverse physiological roles and different pharmacological properties. Early investigations revealed distinct classes of Ca2+ currents which were identified with an alphabetical nomenclature [5]. P/Q-type, N-type, and R-type Ca2+ currents are observed primarily in neurons, require strong depolarization for activation [6], and are blocked by specific polypeptide toxins from snail and spider venoms [7]. P/Q-type and N-type Ca2+ currents initiate neurotransmitter release at most fast synapses [1,8,9]. The Ca2+ channels are composed of four or five distinct subunits (Figure 1a) [8,10]. The α1 subunit incorporates the conduction pore, the voltage sensors and gating apparatus, and target sites of toxins and intracellular regulators. The α1 subunit is composed of about 2000 amino acid residues and is organized in four homologous domains (I–IV) (Figure 1b). Each domain consists of six transmembrane α helices (S1 through S6) and a membrane-associated P loop between S5 and S6. The S1 through S4 segments serve as the voltage sensor module, whereas transmembrane segments S5 and S6 in each domain and the P loop between them form the pore module [11]. The intracellular segments serve as a signaling platform for Ca2+-dependent regulation of neurotransmission, as discussed below.
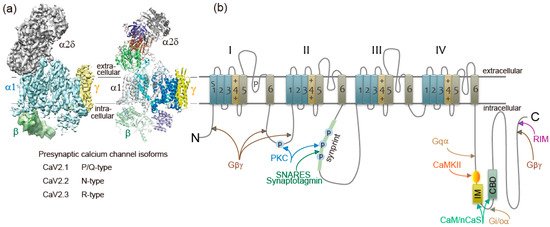
Ca2+ channel α1 subunits are encoded by ten distinct genes in mammals, which are divided into three subfamilies by sequence similarity [2,8,13]. The CaV2 subfamily members CaV2.1, CaV2.2, and CaV2.3 channels conduct P/Q-type, N-type, and R-type Ca2+ currents, respectively [2,8,9,13].
CaV channels are complexes of a pore-forming α1 subunit and auxiliary subunits. Skeletal muscle CaV channels have three distinct auxiliary protein subunits [8] (Figure 1a), the intracellular β subunit, the disulfide-linked α2δ subunit complex, and the γ subunit having four transmembrane segments. In contrast, brain neuron CaV2 channels are composed of the pore-forming α1 and the auxiliary β subunit [14]. The auxiliary subunits of Ca2+ channels have an important influence on their function [15,16]. The CaVβ subunit shifts their kinetics and voltage dependence of activation and inactivation [15,16]. Cell surface expression of the α1 subunits is enhanced by the CaVβ subunit [15,16]. The α2δ subunits are potent modulators of synaptic transmission. The α2δ subunits increase not only Cav1.2 but also Cav2.2, Cav2.1 currents, suggesting that the α2δ subunits enhance trafficking of the CaV channel complex [17]. Expression of α2δ subunits also appears to play a role in setting release probability [18]. Further details of these regulatory interactions are discussed below.
3. Intracellular Molecules Modulate Presynaptic Ca2+ Channels Activity
3.1. G Proteins
3.2. Active Zone Proteins
3.3. t-SNAREs and Synaptotagmin-1
3.4. Ca2+-Sensor Proteins
Ca2+ elevation regulates CaV2.1 channels activity by its binding to CaM [8,78,79,80,81] and related neuron-specific Ca2+-binding proteins, CaBP1, VILIP-2 [82,83,84], and NCS-1 (frequenin) [85]. The presynaptic CaV2.1 channel proteins consist of a pore-forming α1 subunit associated with β, and possibly α2δ subunits (Figure 1a) [86]. The intracellular C terminus of the α1 subunit [81] called the IQ-like motif, which begins with the sequence isoleucine-methionine (IM) instead of isoleucine-glutamine (IQ), and the nearby downstream CaM-binding domain (CBD) are the interacting sites with these Ca2+-binding proteins (Figure 1b). Displacement with alanine in the IQ-like domain inhibited Ca2+-dependent CaV2.1 channels facilitation [78,81], whereas deletion of CBD inhibited Ca2+-dependent CaV2.1 channels inactivation [79,80,81,83,84]. Ca2+/CaM-dependent inactivation of CaV2.1 channels, dependent on global elevations of Ca2+, is observed in transfected cells overexpressing CaV2.1 channels [78,79,80] and in the nerve terminals of the calyx of Held [87,88] where CaV2.1 channels are densely localized. In contrast, the large neuronal cell bodies of Purkinje neurons [89] or SCG neurons [90] rarely show Ca2+-dependent CaV2.1 channels inactivation.
4. Synchronous Neurotransmitter Release Regulated by Ca2+ Channel/SNARE Proteins Complex
Synprint peptides derived from CaV2.2 channels reduced transmitter release from the microinjected presynaptic SCG neurons in culture, due to competitive uncoupling of the endogenous Ca2+ channel-SNARE proteins interaction in nerve terminals [99]. Synprint peptides selectively inhibited fast synchronous synaptic transmission, while they increased late asynchronous release (Figure 3b). Similarly, synprint peptides reduced transmitter release from embryonic Xenopus spinal neurons [100]. Increasing the external Ca2+ concentration effectively rescued this inhibition, implying that synprint peptides competitively displaces docked SVs away from Ca2+ channels, and this effect can be overcome by increasing Ca2+ influx into presynaptic terminals [100].
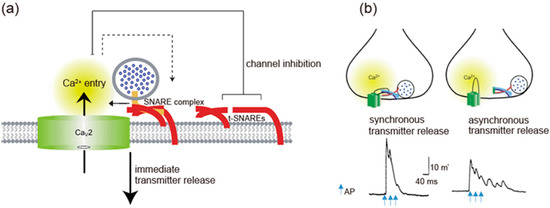
At the calyx of Held, presynaptic neurons express P/Q-, N- and R-type Ca2+ currents in postnatal day 7 rats. P/Q-type Ca2+ currents are more effective than N-type Ca2+ currents and R-type Ca2+ currents in eliciting neurotransmitter release [101,102,103]. The high efficiency of P/Q-type Ca2+ currents to initiate neurotransmitter release is correlated with the close localization of CaV2.1 channels near docked SVs [104], as shown by immunocytochemistry [105], suggesting localization of CaV2 channels determines the efficiency of neurotransmitter release in response to neural activity.
CaV2 channels interaction with SNARE proteins, that is dependent on Ca2+ concentration [63], have two opposing effects: at the pre-firing state synaptic transmission is blocked by enhancing CaV2 channels inactivation, whereas immediately after AP firing tethering SVs near the point of Ca2+ entry enhances synaptic transmission. The overexpression of a syntaxin mutant that is unable to regulate CaV2.2 channels, but still binds to them [72], increased the efficiency of synaptic transmission at Xenopus neuromuscular junctions, as reflected in increased quantal content [106]. In contrast, injected synprint peptides reduced the basal efficiency of synaptic transmission, as reflected in reduced quantal content of synaptic transmission [106]. These results demonstrate a bidirectional regulation of synaptic transmission in vivo by interactions of CaV2.2 channels with SNARE proteins.
This entry is adapted from the peer-reviewed paper 10.3390/ijms20092217