Morphological and anatomical traits of the Vinca leaf were examined using microscopy techniques. Outdoor Vinca minor and V. herbacea plants and greenhouse cultivated V. major and V. major var. variegata plants had interspecific variations. All Vinca species leaves are hypostomatic. However, except for V. minor leaf, few stomata were also present on the upper epidermis. V. minor leaf had the highest stomatal index and V. major had the lowest, while the distribution of trichomes on the upper epidermis was species-specific. Differentiated palisade and spongy parenchyma tissues were present in all Vinca species’ leaves. However, V. minor and V. herbacea leaves had a more organized anatomical aspect, compared to V. major and V. major var. variegata leaves. Additionally, as a novelty, the cellular to intercellular space ratio of the Vinca leaf’s mesophyll was revealed herein with the help of computational analysis.
- Vinca leaves
- parenchymal tissues
- ultrastructure
- electron microscopy
- epicuticular waxes
- cuticle
- computational analysis
- mesophyll airspace
1. Introduction
The Vinca genus belongs to the Apocynaceae family and comprises three species and one variety: V. major L. (bigleaf periwinkle), V. minor L. (lesser periwinkle), V. herbacea Walds. & Kit., (herbaceous periwinkle), and V. major L. var. variegata ‘Louden’ (greater periwinkle with white margins) [1]. Other databases complete the list of species with Vinca difformis Pourr., Vinca erecta Regel & Schmalh., and Vinca ispartensis Koyuncu & Eksi [2].
Vinca plants are intensely studied for their medical properties [3,4,5] due to the rich alkaloid content [6,7,8,9]. These natural products are produced and stored in the aerial parts of Vinca plants [6,10,11], serving as protection against herbivores and pathogens. [12,13]. V. minor is one of the most important medicinal species [14] and the sole source of vincamine in nature, one of the few alkaloids with beneficial effects on cells [3,15]. Quantitative and qualitative differences of the chemical composition in Vinca extracts are dependent on species and the high content of alkaloids, flavonoids, and phenolic compounds, are in correlation with different pharmacological effects [3,16,17,18,19].
In extract preparation, besides the plant species, many factors influence the variability and diversity of the chemical composition, such as the plant organ and tissue, harvesting period and environmental conditions, or extraction method and the solvent used [3,6,7,8]. Previous results confirmed that the chemical composition varies with the Vinca extract, obtained from different organs such as the leaves of V. minor [6,7,14,20]; flowers, leaves, and/or roots of Vinca sardoa [21]; or from other different species of Vinca [4,8].
Undoubtedly, the leaves are very important organs in this matter since other studies showed their direct connection with the alkaloid content. Plant natural products serve in defence mechanisms and are a mixture of substances that often include alkaloids. In Euphorbia characias the aqueous latex produced in the leaves is combined with polyphenols and alkaloids [22]. In Nicotiana stocktonii [23] and Thymus quinquecostatus [24], alkaloids are stored in the glandular trichomes found on the surface of the leaves. In Ficus carica, the alkaloids are synthesized in the epidermal cells and secreted on the surface of the leaves by the glandular trichomes [25,26].
Based on similar studies as the aforementioned, the leaves of Vinca species are often used for extract preparation due to the fast release of high alkaloid concentration [11,27], but to our knowledge, their morphology and anatomy are insufficiently studied highlighting the scope of this study. The chemical composition of Vinca extracts varies to a great extent due to plant photosynthesis [28,29], which is regulated by numerous environmental and internal factors. These factors include morphological and anatomical leaf traits, i.e., shape, size, and the number of stomata, type or number of trichomes, and the weight of epidermal cuticle [30,31].
2. Vinca Leaf Epidermises
The epidermises of leaves were studied by scanning electron microscopy (SEM) and details such as the presence or absence, type, shape, and distribution of trichomes and stomata were observed.
Pavement cells on the adaxial side of V. minor, V. major, and V. major var. variegata leaves had similar typical jigsaw puzzle shapes (Figure 1a,c,e), whereas pavement cells of V. herbacea were papillary shaped (Figure 1g). Except for V. minor, all species had stomata on the upper epidermis. Tector trichomes with various ornamentations were present on the adaxial leaf side of the leaves as well (Figure 1b,d,f,h).
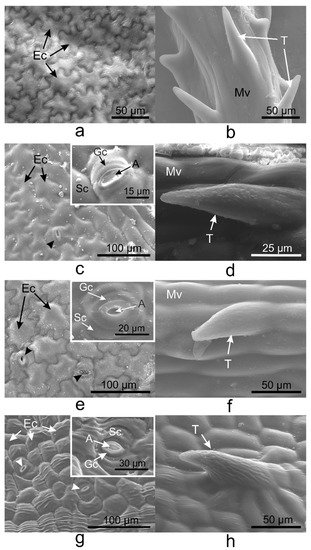
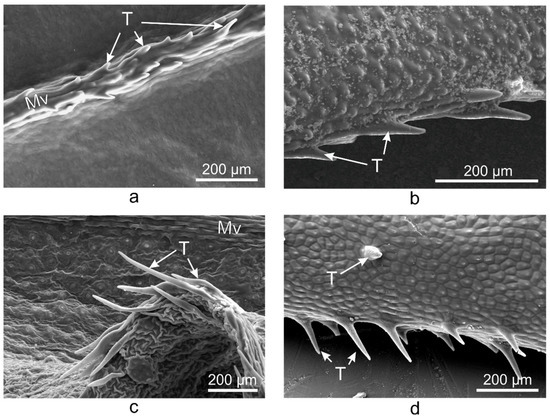
On the abaxial leaf side, all Vinca species presented typical puzzle-shaped pavement cells, while stomata were randomly distributed on the entire surface, except for the midvein (Figure 2).

Figure 2. SEM micrographs showing the abaxial (lower) epidermis of the Vinca leaves with an overview along the midvein and detailed view of the stomata. (a) V. minor, (b) V. major, (c) V. major var. variegata, (d) V. herbacea; A—aperture, Ec—epidermal cell, Gc—guard cell, Mv—midvein, Sc—subsidiary cell, arrowheads—stomata.
3. Vinca Trichomes and Stomata Characteristics
| Species | SD * | SI (%) * | SL (µm) | SPI (%) | TD * | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ad | Ab | Ad | Ab | Ad | Ab | Ad | Ab | Ad | |
| V. minor | 0 | 223 | 0 | 25.3 | 0 | 9.57 ± 0.6 | 0 | 2.04 | 31 |
| V. major | 10 | 49 | 4.5 | 23.2 | 14.84 ± 1.1 | 19.75 ± 0.4 | 0.22 | 1.91 | 14 |
| V. major var. variegata | 12 | 68 | 4.2 | 20.9 | 12.33 ± 1 | 12.24 ± 1.4 | 0.18 | 1.01 | 25 |
| V. herbacea | 16 | 60 | 4.4 | 25 | 12.56 ± 1 | 13.05 ± 1.2 | 0.25 | 1.02 | 23 |
4. Vinca Mesophyll
| Species | LT (µm) | PTT (µm) | STT (µm) | CTR (%) | SR (%) |
|---|---|---|---|---|---|
| V. minor | 85.47 ± 2.6 | 12.58 ± 0.7 | 52 ± 0.5 | 14.72 | 60.84 |
| V. major | 136.85 ± 1.4 | 0 | 124 ± 0.5 | 0 | 90.61 |
| V. major var. variegata | 97.09 ± 1.5 | 14.57 ± 4.4 | 67.29 ± 1.7 | 15.01 | 69.31 |
| V. herbacea | 133.38 ± 2.2 | 36.66 ± 2.2 | 68.42 ± 3.7 | 27.48 | 51.3 |
5. Discussion
There are quantitative and qualitative differences regarding the chemical composition of Vinca extracts, and previous studies showed that the morphological and anatomical traits of the leaves (shape and surface, cuticle, stomata, trichome, epidermis, etc.) are responsible for the chemical composition variability in the extracts [33,34,35,36]. These morphological and anatomical aspects of the Vinca leaf had been investigated in this study as well. The leaf surface area and dry leaf mass are directly correlated for vines species (Vinca major, Trachelospermum jasminoides, Hedera nepalensis var. sinensis) [37] and for Arabidopsis thaliana [38]. This aspect is important in extract preparation for maximal exploitation of the plants.
Additionally, the morphological characters of all plant organs are used for accurate identification of species [14,33,34,35]. The leaf shape varies within species and populations, and even within the same individual plants; therefore, it affects photosynthesis, water balance, temperature control, and the interactions with other organisms [39].
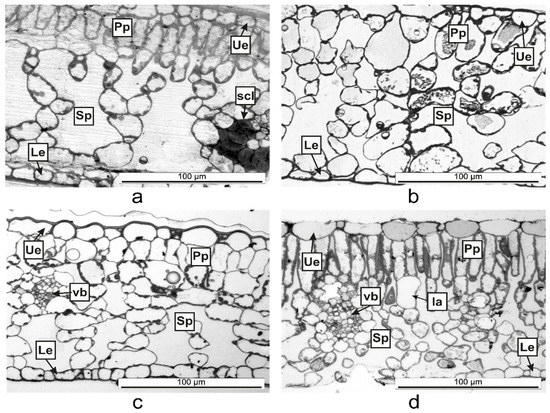
On the same surface of the Vinca leaf within different species, the epidermal cells are distinct, and here was shown that they could vary on both sides even for the same species (Figure 1, Figure 2 and Figure 3). The Vinca leaf epidermis has a single layer of cells composed of pavement cells, stomatal complexes, and trichomes (Figure 1, Figure 2, Figure 3, Figure 4 and Figure 5), that serves as a protective barrier to environmental factors [40]. On the adaxial side, Vinca leaves had puzzle-shaped epidermal cells, except for V. herbacea that had papillary epidermal cells. Similar results were previously reported for V. minor and V. herbacea only [41], and there are several theories that try to explain this type of organization of the pavement cells [42,43,44].
These specialized epidermal cells differ intra- and interspecific, depending on various extrinsic and intrinsic parameters, e.g., the leaf age, dimension, insertion position on the stem, combined with CO2 atmospheric concentration, irradiance, humidity, and other environmental factors (pollution, pathogens, etc.) [44,45,46]. Therefore, the current study was conducted under minimum variation of these parameters. The outdoor plants (V. minor and V. herbacea) grew in a shaded area and had the same light exposition (46°45′51” N; 23°34′47” E). The indoor plants (V. major and V. major var. variegata) were exposed only to sunlight with the same circadian cycles as the outdoor plants and the humidity and CO2 levels were regulated so that they could resemble the natural conditions. All the above-mentioned criteria play vital roles for the medicinal taxa [47,48].
Also, a connection between the cell walls of the leaves’ epidermis and the natural products in plants had been previously established [6]. In most plant species, the epidermis has specialized roles in the biosynthesis and accumulation of a wide range of natural products, including alkaloids [40], terpenes, and flavonoids [40,49], indicating that the variances observed here could be indicative of the chemical variation of the plant extracts.
The Vinca leaf epidermal cells are covered by a cuticle (Figure 5, Figure 7 and Figure 8) with complex chemical composition and have different thicknesses on the upper and lower epidermises as previously indicated [50,51,52]. In terms of electron-dense cutin layers, the V. minor, V. major var. variegata and V. herbacea leaves are similar to the leaves of Pyrus communis and Populus bolleana [50], but were never described for the Vinca species. The cuticular waxes play significant roles in Vinca species due to their involvement in the defense mechanisms against pathogens, temperature variations, salinity, or excessive ultraviolet radiations [52].
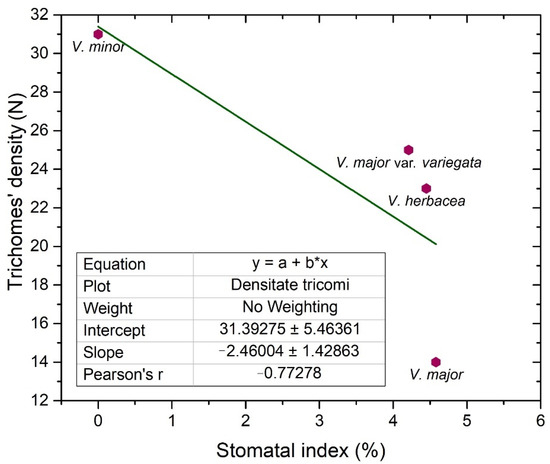
Glandular and secretory trichomes are responsible for biosynthesis, secretion and/or accumulation of phytochemicals useful for defense [40]. In the leaves of Vinca species analyzed herein, only tector (protective) trichomes had been identified, with the main role in defense mechanisms against water loss. The number of trichomes increases while the stomatal index decreases, and this way, the transpiration process is balanced [53], explaining our results where the stomatal index was found to be inversely proportional to trichomes’ density. For V. minor, the trichomes were placed only on the midvein, while the other species had trichomes on the margins and on secondary veins. Similar results had been previously reported as well for V. minor and V. herbacea [41,54].
Stomata are important structures involved in exchange processes of the plant with the environment, balancing the CO2 influx during photosynthesis and water vapor efflux during transpiration [28,33,35,40,55]. Stomata exhibit a diverse range of shapes, sizes, and numbers across different plant species [53]. Leaf stomata in plants from the same genus or from different plants of the same species are variable [56,57]. Significant negative correlation was documented between stomatal density and stomatal shape parameters (stomatal area, stomatal perimeter, stomatal long axis, stomatal short axis) in other plant species as well [58].
The stomatal index is the recommended parameter to be measured since it reports the stomata density in relation to the number and size of the pavement cells, in each area [28,46]. The stomatal index for the adaxial side (around 4%), is evidently lower than the one registered for the abaxial side (20–30%); therefore, the studied Vinca species are all hypostomatic.
Our results showed that the overall thickness of the Vinca leaf differs with the species and this influences the photosynthetic rates as well [59]. Besides this, the stomatal index has different values for each Vinca species, and the chemical composition could vary as well. This is because a higher specific stomatal index is related to a higher net rate of photosynthesis, and consequently, the plant could enhance the carbon fixation [28,32]. For example, several sugars were shown to affect the growth and development of V. minor and C. roseus leaves [14]. As carbon is present in many sugars and alkaloids, this could also influence the chemical composition in the long run [28].
Vinca leaf mesophyll has different thicknesses, a parameter which is species depended [36]. The Vinca leaves contain different numbers of columnar palisade cells. It was confirmed that sunlight-grown plants have more columnar palisade cells than those of shade-grown plants [29,34,35]. Similar to our results where the indoor plants had a less differentiated palisade tissue than the outdoor plants, other studies showed that the palisade tissue cells in V. major var. variegata leaves were single-layered for the plants located at roof level and double-layered for ground-level plants [35]. Another study demonstrated that depending on light exposition, the leaf of V. minor had a double-layered palisade tissue and V. herbacea leaf, a single-layered one [41]. The shape of palisade cells and the movement of chloroplast in accordance with the light conditions, are essential for efficient leaf photosynthesis, by cell development regulation with the help of phototropin [29]. The number and shape of palisade cells, which compose the photosynthesis unit area, are different for each Vinca leaf analyzed herein, therefore the photosynthesis process varies along with the anatomical traits [29] and this might influence the chemical composition as well if the plants are to be considered for extract preparation.
The thickness of the spongy parenchyma in the Vinca leaf varies with the species and contains several layers of ovoid, oblong, or circular shaped cells and the mesophyll airspace [34,35,41]. The cells of the spongy parenchyma store important nutrients for the plant [60]; therefore, its thickness would be directly related to the abundance of natural compounds [61]. Other studies described 7 to 8 layers of cells in the spongy tissues of V. minor and V. herbacea [34,41], while for V. major var. variegata, the number of layers decreased to 6–7 for plants grown at ground level [35]. The results presented herein showed that the plants grown outdoor had 7 to 8 layers, while the plants grown indoor, had 5 to 6 layers of cells in the spongy parenchyma—results which are consistent with other findings—and this indicates that growth conditions are important parameters that should be considered before extract preparation.
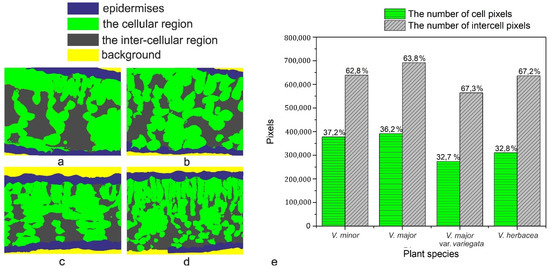
The programmatic method used for computation helped to determine the ratio between intercellular spaces and cells in the mesophyll of the Vinca species. First, it must be stressed that the main objective was the accuracy of the mathematical computations with respect to the ground truth. Initially, various methods were employed for the semi-automatic segmentation of the regions in the semithin sections, using state-of-the-art methods such as those based on computer vision [62,63] or machine learning approaches [64,65]. The employed methods obtained the same accuracy levels as those found in the literature.
However, the visual assessment of the results leads to the conclusion that since the end goal was to compute a percentage as close to reality as possible through an automated method of visual segmentation and labeling of the cellular and inter-cellular regions, significant errors in the computational process occurred. In other words, an accuracy level that is acceptable from a computer science perspective may, however, damage the biological scientific accuracy of the research. V. minor had the lowest mesophyll airspace (62.8%), followed by V. major (63.8%) and very close to each other were V. major var. variegata and V. herbacea (67.3% and 67.2%, respectively). According to recent studies, there is a connection between the mesophyll airspace and functional stomata [36,66], implying that larger airspaces are connected to a higher stomatal index. Since the Vinca species are hypostomatic, the high percentage for intercellular spaces is justified in relation to the high stomatal index observed.
The ultrastructural analysis of the leaves offered a more detailed view of the anatomy and confirmed some of the hypotheses regarding the spongy parenchyma. Lipids are used as substitutes for energy production and their presence as big droplets in the V. major and V. major var. variegata leaf could indicate a compensatory mechanism adopted by the plant because of the greenhouse storage conditions [67]. As indicated in other studies, alkaloids are harmful to the plant and are therefore stored or synthesized in small amounts [61] and only when needed, and the chemical composition variability of Vinca plant extracts could be reflected by different amounts of lipid droplets identified in leaf spongy parenchyma [67]. Overall, the methods used to determine the morphological and anatomical aspects of the Vinca leaves are correlated with each other and can be used to analyze the leaves of other species as well. This is an important step since Vinca species are recognized for their medicinal properties, and a brief morphological and anatomical description of a batch could help determine if the plants are suited for extraction.
This entry is adapted from the peer-reviewed paper 10.3390/plants10040622