Charles Darwin used domestication as a metaphor for natural selection because everyone is familiar with the term. Since Darwin, domestication has become a major topic of research, both to help understand evolution, as Darwin did, and to understand how human societies came to dominate the world, which also interested Darwin. However, Darwin did not define the term, which has allowed students of domestication, such as geneticists, archaeologists and others, to tailor their definitions to their own objectives. Even our grammar influences the way we interpret simple phrases about domestication ( p. xiv), as we tend to put ourselves, as individuals or the human collective, in charge. As Darwin pointed out, however, the long history of human interaction with plants and animals that resulted in the domestication of some of them was influenced more by unconscious selection than conscious selection. Only when considering the latter type of selection, which Darwin called methodical selection, can we affirm that humans are in charge, and even then unintended consequences are common. Nonetheless, especially when used in studies to understand how humans came to dominate the world, definitions often assume that humans are in control, which is unlikely at the beginning of human interactions with plants and animals.
As pointed out by Dolores Piperno and Deborah Pearsall ( p. 6), clear definitions of concepts are extremely important when discussing domestication and the food production systems in which many domesticates are grown or raised. Melinda Zeder reviewed a set of definitions to highlight their differences and theoretical frameworks, and emphasized that definitions for plants and animals are often quite different. Clement et al. prepared a short list of definitions for plant domestication since Zeder. Full definitions can help identify nuances that place human culture within Nature, as occurs in most Neotropical ontologies, rather than outside, which is typical of ideas of being in charge. We think that returning to a dictionary is also an appropriate exercise to explore definitions, especially as the definitions in Zeder and Clement et al. were designed by their authors to meet their own objectives, the majority of which are associated with identifying changes in morphology or genetics that prove that human selection resulted in a response.
We will use the 1989 edition of the Oxford English Dictionary (OED), whose first edition was being researched and published at the same time that Darwin was writing. The verb domesticate comes from the Latin domesticäre – to dwell in a house, to accustom. The house is the center of the domus, the Latin root of domesticäre. The OED definitions include: “1.a. To make, or settle as, a member of a household; to cause to be at home; to naturalize. 1.b. To make to be or to feel ‘at home’; to familiarize. 2. To make domestic; to attach to home and its duties. 3. To accustom (an animal) to live under the care and near the habitations of man; to tame or bring under control; to civilize. 4. To live familiarly or at home (with); to take up one's abode.” It follows that the noun domestication is “the action of domesticating, or the condition of being domesticated” (OED), i.e., both the process itself and the results of the process.
These definitions are about humans, who domesticate each other in their houses, with their associated gardens, orchards, pastures, woodlots, agroforests, and adjacent managed forests. Hence, the house and the surrounding landscape comprise the domus , until humans started living in cities or more recently in apartments. Animals are recognized in one of the definitions as being domesticated in the domus also, and it was Darwin who included plants. From these definitions, domestication is clearly anthropocentric, i.e., it is about us humans. Note that only one definition mentions control.
What are the human behaviors involved? Two are explicit in the definitions: care (of the occupants of the domus) and duty (the tasks of caring for the domus and its occupants). Two are implicit. Selection – since humans are selective about what is brought into the domus. The fact that selection is implicit, rather than explicit, may be why Darwin adopted domestication as his metaphor for natural selection. Accumulation – as people and animals are brought into the domus, both from nearby (familiarize) and far away (naturalize). As Darwin recognized, humans like variety, which can be seen in most gardens and even more clearly in swiddens. The definitions above are about both organisms (humans, animals, plants) and the domus. Most current definitions of domestication concentrate one behavior (selection), but care and accumulation are just as important.
- landscape domestication
- Mesoamerica
- plant domestication
- plant management
Domestication in the Neotropics
1. Domestication as Process
According to David Rindos [1], domestication is a co-evolutionary process that involves relationships among humans and other organisms. He divided the process into three stages, while recognizing that the division is somewhat arbitrary and used for convenience: (1) incidental domestication, which includes simple dispersal and protection actions by people, creating and maintaining coevolutionary interactions outside agroecological systems; (2) specialized domestication, which involves forces that initiate and maintain the agroecological system; and (3) agricultural domestication, which involves forces controlling the function, evolution, and spread of developed agricultural systems ([1] p. 153). In this perspective, selection and caring, in the form of simple dispersal and protection, are at the beginning of the process, while an agroecological relationship originates where humans create conditions for the growth of useful plants, or what Piperno and Pearsall ([2] p. 6) call cultivation. The creation of an agroecological system starts in a new settlement, especially with its associated dump heaps, and gradually extends outwards. As Rindos points out, this creates new ecological conditions, or niches, and some plants take advantage of them. These colonizing species, also called weeds, benefit from the processes that create agroecologies and some of these species are useful to humans, so they are maintained and protected [3][4][5]; they may be selected and become domesticated [5][6].
In the Neotropics, the process started with the arrival of humans about 30,000 years ago. Upon arrival they identified useful species and the variation within each, allowing the preliminary selection of individuals for gathering. The gathering starts dispersal towards the settlement ([1] pp. 112-120), as some fruit or seeds may fall en route, or may be consumed and the seeds excreted en route [7][8]. The best individuals in the population (and later those dispersed en route) are protected, others may be tolerated, the worst may be eliminated if they compete with preferred individuals or good individuals of other useful species [7]. All of these activities gradually result in a population more useful to humans in the original ecosystem, which is gradually becoming a domesticated landscape—all without creating an agroecology, hence Rindos’ term incidental domestication ([1] pp. 154–158).
As mentioned, the agroecological system, with its specialized domestication ([1] pp. 158–164), appears in human settlements, all of which have dump heaps. The dump heap origin of crop domestication and food production has a long history [9]. What is most important is that dump heaps and nearby areas of the settlement develop into gardens [10][9], which are recognizable agroecological systems that can be replicated beyond the settlement, giving rise to more intensive agroecological systems where agricultural domestication continues ([1]pp. 164–166). The latter is what most people consider to be domestication, as seen in the standard narrative of crop domestication and agricultural origins, e.g. Lien et al. [11]. By recognizing only the latter stage, a long history of human–plant interactions is ignored.
The concept of domesticated landscapes has been used for a century (see Smith [12] for a list of synonyms) and can conceptually be disentangled from domestication of plants and animals [13]. The concept can be defined as a process by which human manipulation of the demography of plant and animal populations changes the landscape’s ecology, resulting in a landscape more productive and congenial for humans [13][14]. At its simplest, the protection and dispersal involved in incidental domestication promote the initial changes ([1] pp. 112–120). As management intensity increases, with the removal of competitors, intentional planting of seeds and seedlings, mulching around these, and other practices to care for individual plants, the landscape becomes more productive [3][4][7][15][16]. A continuum of change can be evident, as humans invest more effort in caring for some useful plants. These changes are important for both humans and plants, and create conditions favorable for other species of plants and animals as well [17].
Cultivation introduces more dramatic changes in the ecosystem, as noted by Rindos[1]. Although cutting trees comes to mind first, William Denevan [18] pointed out that with a stone axe it is easier to find a clearing where a large tree fell or a wind storm had opened a larger area. Once open, fire becomes an essential tool [18][19] ([20] pp. 37–43). All the practices already mentioned are used to propagate and care for useful plants in a new agroecosystem. Today this generally starts as a horticultural plot and turns into an agroforestry plot, often seen as mimicking natural ecosystems, especially as local species volunteer and are tolerated in the agroecosystem [21].
2. Domestication as Result (the Domestication Syndrome)
“Since domestication is an evolutionary process, there will be found all degrees of plant and animal associations with man and a range of morphological differentiations from forms identical to wild races to fully domesticated races. A fully domesticated plant or animal is completely dependent upon man for survival.” ([6] p. 62). This observation calls for a definition of a domestication continuum, rather than using a general definition that does not discriminate any possibilities along the way except the last one, when the domestication syndrome is clearly visible. As Darwin observed, humans select – unconsciously or more rarely consciously – for a small number of traits that make their selected plants different from wild ones [22]; this set of traits is the domestication syndrome [23], which varies in composition among the different species that humans manage and cultivate.
A definition that recognizes a continuum, following from Rindos, is that plant domestication is a co-evolutionary process during which human selection of the phenotypes of wild, promoted, managed or cultivated plants results in changes in the next generation’s phenotypes and genotypes that make them more useful to humans and better adapted to domesticated landscapes [14]. According to Harlan, during the beginning of the process the changes are so subtle that they are hard to differentiate from wild populations ([6] p. 64), with the clear implication that the domestication syndrome contains only incipient changes in one or a few traits, and may only be visible as reduced variability [4][14].
The results of the domestication continuum extend from the first incipient changes to a clearly differentiated domestication syndrome [14], and are the major interest of the majority of the definitions about this process [24][25]. Along the continuum, some categories can be identified (Figure 1), mostly for convenience in discussing the concept [14]. An incipiently domesticated population has both the mean and variance of a selected trait within the variation of the species ([6] p. 64). A semi-domesticated population has more pronounced differentiation, while a domesticated population may extrapolate the variation of the wild populations and also has become dependent on humans (the last stage mentioned by Harlan). Although this sequence is defined as being linear, the world is much more complex. There are multiple ways to get from A to D, and there is no guarantee that C or D will be attained or, if attained, maintained through time.
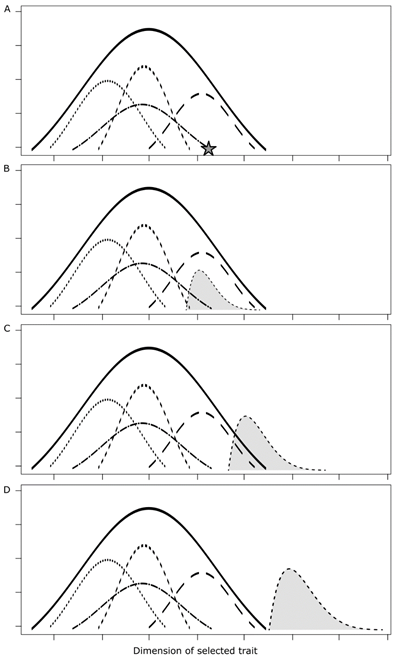
Figure 1. Hypothetical domestication continuum (frequency distributions for population means and variances of the dimension of a phenotypic trait of interest to humans, e.g., fruit size). (A) A species with four wild populations. A few plants are selected in one population (star) to create a new population in (B). (B) As above, with one incipiently domesticated population. Observe that the variance of the incipient domesticate is smaller than that of the wild populations, due to the selection of a small number of plants to create the new population—a result of the founder event. Observe also that the mean and variance are within the variance of the species. (C) As above, with one semi-domesticated population with somewhat greater variance. (D) As above, with one domesticated population. Observe that both the mean and the variance are outside of the variance of the species. Observe in B–D that the domesticated populations have skewed distributions, with more variation towards the right side, representing directional selection, e.g., for larger fruit. Adapted from Leakey et al. [26], with thanks to Alessandro Alves-Pereira.
During the mid-20th century most students of domestication thought that the process could be quite rapid, e.g., from wild to domesticated phenotypes, such as the non-shattering seed rachis of wheat (Triticum spp.) or barley (Hordeum vulgare), in 200 years [6]. This fitted nicely with the standard narrative about the rise of states [20]. Since the turn of the millennium, archaeologists found that the wheat and barley domestication processes took thousands of years [20][27], and that “pre-domestication cultivation” occurred in south-western Asia in the late Pleistocene [28], similar to the situation for “low-level food production” in the Americas[29]. At the same time, geneticists used the biological model of domestication to identify how humans interact with plant genetics. The biological model has two interacting equations [30]:
(1) VP = VG + VE + VGxE
(2) R = h² × i × √VP
Equation (1) explains the relationship among variances of phenotypes (VP), genotypes (VG), environment (VE) and the genotype-by-environment interaction (VGxE). As in Figure 1, these variances are of any trait of interest to humans in a population, e.g., fruit size. This equation is about what is available to humans (VP) and also explains how phenotypes are created during growth depending upon genotypes and their interactions with their environment. Equation (2) explains the response (R) when humans select (i) from the population. The narrow sense heritability (h2) is that proportion of VG that explains the similarity between parents and offspring for the trait of interest [30]. The greater h2, i, VP, the greater the response; reduce any variable and the response decreases. Each trait of the domestication syndrome can be analyzed this way.
The model used by geneticists found that the intensity of selection for the non-shattering rachis of wheat and barley was extremely low, only slightly different from that expected from natural selection, and that it took thousands of years for the non-shattering trait to reach even low proportions in the pre-domestication cultivated populations [31][32][33]. These authors did not use the term, but this is what we mean by incipient domestication, a change so small that it is hard for archaeologists to identify.
Most geneticists and many archaeologists who study domestication do not give much attention to an extremely important component of the model: VE. Rindos[1], however, was very clear that plant domestication occurs within domesticated landscapes and that, as the human investment in agroecological management expands, so does the response to selection. Equation (1) states that VP is the sum of three other variances and can be modified by changes in any of them. Like VG, which contains several genetic components, VE contains numerous biotic and abiotic components typical of niches [34], such as soils, water availability, pollinators, pests, diseases, herbivores. Humans and their management practices are also biotic and social components of the niche, which becomes an agroecological niche as human action increases. Choices about where to plant, when to plant, how to fertilize, irrigate, weed, etc., all affect VE and feed into VP, both directly and via VGxE [35]. Since these can change VP without human selection (i), it is possible to obtain a response (R) without human selection, so management of the agroecology is always an important consideration. Observe also that all these management options are designed to meet the needs of plants who respond to this care, e.g., this response is plant agency [1]. When there is human selection, management practices enhance the response.
3. Food Production Systems
The term food production systems represents the agroecology described by Rindos [1] and one of its many variants is included explicitly or implicitly in the majority of definitions used by geneticists and archaeologists [24][25]. There are numerous types of food production systems [36], each with somewhat different agroecosystems and human decisions about crops and their management. The two most commonly used terms are horticulture and agriculture. Horticulture comes from the Latin hortus (garden) and cultūra (culture or cultivation), and is defined as “The cultivation of a garden; the art or science of cultivating or managing gardens, including the growing of flowers, fruits, and vegetables” (OED). Fruits are often produced by trees, which have their own term: arboriculture (from the Latin arbor—tree). Since many fruits and other products are also produced on plants that are not trees (e.g., palms, cacti, agaves) and which occur in some types of forest, it is also appropriate to define silviculture (from the Latin silva—forests or stands of trees). Similarly, agriculture comes from agrī (genitive of ager—field) and is defined as “(a) Originally: the theory or practice of cultivating the soil to produce crops; an instance of this (now rare). (b) Later also (now chiefly): the practice of growing crops, rearing livestock, and producing animal products (as milk and eggs), regarded as a single sphere of activity; farming, husbandry; (also) the theory of this”, (OED). In modern usage, agriculture is thus all inclusive, but its original use was for crops, which are defined as “The annual produce of plants cultivated or preserved for food, esp. that of the cereals; the produce of the land, either while growing or when gathered; harvest”, (OED). This is why agriculture is generally associated with southwestern Asia, where wheat and barley were domesticated in fields, although gardens, including with cereals ([20] p. 43), other annuals and perennials, were certainly earlier than fields, although seldom emphasized (e.g., [6]).
Agriculture is the term of choice in the standard narrative about the rise of states. In this narrative, horticulture is small-scale (gardens), even “primitive”, compared to agriculture, which is large-scale (fields), with advanced technologies, such as draft animals to pull plows and operate threshing equipment to separate chaff from grain, etc. Scale often decides the usage [41]. A recent article about the expansion of maize (Zea mays) use and production in pre-Columbian Mexico asks “Is it agriculture yet?” [37], referring directly to the scale of use and production. A majority of scholars follow this usage, e.g., [2].
Since we are discussing domestication, however, another factor is important. Oake Ames [38], observed that in horticulture plants are treated as individuals, while in agriculture plants are treated as groups (or populations). This is an extremely important observation because it has to do with selection (i) and thus response to selection (R) in the biological model. Those of us with gardens often talk or sing to our plants and they respond to our care, especially if we weed, fertilize and irrigate. Anthropologists have reported this among indigenous peoples across the Neotropics [39][40], where local ontologies consider other living beings to be social organisms similar to humans in many respects [41]. In Amazonia, indigenous women consider their manioc (Manihot esculenta) plants to be their children and sing to them to encourage them [42]; in Mexico, the Mixtec, Nahua and other peoples pray in their milpas to encourage and safeguard the maize while they care for the plants[43]. This caring also implies a duty to care for and protect. Although there is an effect on VE, due to the weeding, fertilizing and irrigating, what is more important is selection (i), because the better you know your individual plants the easier it is to decide which ones get more space in the next garden. This is true for plants propagated by seeds (e.g., maize) or vegetatively (e.g., potatoes (Solanum tuberosum) or manioc). The cultivation of manioc and potatoes is a special kind of horticulture, called vegeculture—the culture of vegetatively propagated plants. Harlan ([6] p. 131) observed that vegetative propagation is instant domestication, because the plants depend completely on their humans to be propagated into the next garden.
Another factor is important in the Neotropics: no animals were domesticated that could pull a plow. Hence, all labor in food production systems was human, although ducks may have helped with weeding and pest control [44]. Neotropical societies had numerous tools for working the soil and processing plants, but none that permitted the scale typical of agriculture with draft animals. That is not to say that they produced less food; the early chroniclers marveled at the well-feed, healthy people in the villages and urban areas they conquered [45][46]. In some places, there were moderately large individual fields, such as the raised fields in the Llanos de Mojos, lowland Bolivia, which could be 20 × 50 m, with dozens of such raised fields around some villages [19]. In Central West Mojos, an area of about 10,000 km2, there are about 36,000 raised field platforms, with a total raised-surface area of 100 km2 ([47] p. 105). In the Andes, the tens of thousands of terraces, andenes, each had small surfaces, many much smaller than in Mojos, but summed were able to support the Inca state [19]. Similarly, the milpa and agroforestry systems of the Maya supported its large population [15], and the terraces, chinampas and agroforestry systems of the Aztecs supported another large state [48]. In Amazonia, the chacra horticultural plots and agroforestry systems supported the expansion of the Arawak-speaking peoples [49], and were used by all other ethnic groups that decided to practice horticulture and agroforestry. Importantly, in the more forested Neotropical regions, including the Atlantic Forest and forested savannas in Brazil, these agroforestry systems were complemented by forest management [7][50], and some, perhaps many, societies obtained more food from their forests than from their gardens and agroforests [51][52].
The observant reader will have noticed that an additional term slipped in: agroforestry. Unlike the other terms we have used, this is not derived directly from Latin, but from research on modern small-scale indigenous and traditional food production systems across the tropics [53]. The term suggests a combination of agriculture with forestry. The majority of the hundreds of different agroforestry systems described by PK Nair [53]are in reality combinations of horticulture, vegeculture, arboriculture and silviculture, generally with volunteer plants that are tolerated and may be protected.
Is domestication different in the Neotropics than elsewhere? Yes and no. The mechanics are the same. Humans accumulate variability, care for it, select (both during accumulation and while caring), and through time obtain responses in the form of more variability, larger and tastier fruits and vegetables, more colorful and beautiful flowers, more bioactive compounds (medicinal and magic) etc. The associated food production systems, however, were partially different. There was no agriculture, in the original definition of the term, in the Neotropics before European conquest. The predominance of horticulture, with the focus on individual plants, may explain why there is so much variability.
This entry is adapted from the peer-reviewed paper 10.3390/quat4010004
References
- Rindos, D. The origins of agriculture: an evolutionary perspective; Academic Press: San Diego, 1984; pp. 325.
- Piperno, D.R.; Pearsall, D.M. The origins of agriculture in the lowland Neotropics; Academic Press: San Diego, 1998; pp. 400.
- Casas, A.; Otero-Arnaiz, A.; Pérez-Negrón, E.; Valiente-Banuet, A. In situ management and domestication of plants in Mesoamerica. Ann. Bot. 2007, 100, 1101-1115, doi:10.1093/aob/mcm126.
- Casas, A.; Vázquez, M.d.C.; Viveros, J.L.; Caballero, J. Plant management among the Nahua and the Mixtec in the Balsas River Basin, Mexico: An ethnobotanical approach to the study of plant domestication. Hum. Ecol. 1996, 24, 455-478, doi:10.1007/BF02168862.
- Blancas, J.; Casas, A.; Rangel-Landa, S.; Moreno-Calles, A.; Torres, I.; Pérez-Negrón, E.; Solís, L.; Delgado-Lemus, A.; Parra, F.; Arellanes, Y., et al. Plant management in the Tehuacan-Cuicatlan Valley, Mexico. Econ. Bot. 2010, 64, 287-302, doi:10.1007/s12231-010-9133-0.
- Harlan, J.R. Crops & man, 2 ed.; American Society of Agronomy & Crop Science Society of America: Madison, WI, 1992.
- Levis, C.; Flores, B.M.; Moreira, P.A.; Luize, B.G.; Alves, R.P.; Franco-Moraes, J.; Lins, J.; Konings, E.; Peña-Claros, M.; Bongers, F., et al. How people domesticated Amazonian forests. Front. Ecol. Evol. 2018, 5, 171, doi:10.3389/fevo.2017.00171.
- Parra, F.; Blancas, J.; Casas, A. Landscape management and domestication of Stenocereus pruinosus (Cactaceae) in the Tehuacán Valley: human guided selection and gene flow. J. Ethnobiol. Ethnomed. 2012, 8, 32, doi:10.1186/1746-4269-8-32.
- Anderson, E. Plants, man and life; Courier Dover Publications: Mineola, NY, 2005; pp. 251.
- Hastorf, C. The cultural life of early domestic plant use. Antiquity 1998, 72, 773-782, doi:10.1017/S0003598X00087366.
- Lien, M.E.; Swanson, H.A.; Ween, G.B. Naming the beast--exploring the otherwise. In Domestication gone wild: politicas and practices of multispecies relations, Swanson, H.A., Lien, M.E., Ween, G.B., Eds. Duke University Press: Durham, 2018; pp. 1-30.
- Smith, B.D. General patterns of niche construction and the management of 'wild' plant and animal resources by small-scale pre-industrial societies. Phil. Trans. R. Soc. Lond. B. 2011, 366, 836-848, doi:10.1098/rstb.2010.0253.
- Clement, C.R.; Cassino, M.F. Landscape domestication and archaeology. In Encyclopedia of Global Archaeology, Smith, C., Ed. Springer: New York, 2018; 10.1007/978-3-319-51726-1_817-2pp. 1-8.
- Clement, C.R. 1492 and the loss of Amazonian crop genetic resources. I. The relation between domestication and human population decline. Econ. Bot. 1999, 53, 188-202, doi:10.1007/BF02866498.
- Ford, A. The Maya forest: a domesticated landscape. In The Maya world, Hutson, S.R., Ardren, T., Eds. Routledge: New York, 2020; pp. 519-539.
- Gómez-Pompa, A.; Flores, J.S.; Sosa, V. The “pet kot”: a man-made tropical forest of the Maya. Interciencia 1987, 12, 10-15.
- Bogoni, J.A.; Muniz-Tagliari, M.; Peroni, N.; Peres, C.A. Testing the keystone plant resource role of a flagship subtropical tree species (Araucaria angustifolia) in the Brazilian Atlantic Forest. Ecol. Indicators 2020, 118, 106778, doi:10.1016/j.ecolind.2020.106778.
- Denevan, W.M. Stone vs. metal axes: The ambiguity of shifting cultivation in prehistoric Amazonia. J. Steward Anthro. Soc. 1992, 20, 153-165.
- Denevan, W.M. Cultivated landscapes of Native Amazonia and the Andes; Oxford University Press: Oxford, 2001; pp. 396.
- Scott, J.C. Against the grain: a deep history of the earliest states; Yale University Press: New Haven, 2017; pp. 336.
- Denevan, W.M.; Treacy, J.M.; Alcorn, J.B.; Padoch, C.; Denslow, J.; Paitan, S.F. Indigenous agroforestry in the Peruvian Amazon: Bora Indian management of swidden fallows. Interciencia 1984, 9, 346-357.
- Darwin, C. On the origin of species by means of natural selection, or the preservation of the favoured races in the struggle for life; John Murray: London, 1859.
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and processes in crop domestication: an historical review and quantitative analysis of 203 global food crops. New Phytol. 2012, 196, 29-48, doi:10.1111/j.1469-8137.2012.04253.x.
- Zeder, M.A. Central questions in the domestication of plants and animals. Evol. Anthro. 2006, 15, 105-117, doi:10.1002/evan.20101.
- Clement, C.R.; Casas, A.; Parra-Rondinel, F.A.; Levis, C.; Peroni, N.; Hanazaki, N.; Cortés-Zárraga, L.; Rangel-Landa, S.; Alves, R.P.; Ferreira, M.J., et al. Disentangling domestication from food production systems in the Neotropics. Quaternary 2021, 4, doi:10.3390/quat4010004.
- Leakey, R.R.; Tchoundjeu, Z.; Smith, R.I.; Munro, R.C.; Fondoun, J.-M.; Kengue, J.; Anegbeh, P.O.; Atangana, A.R.; Waruhiu, A.N.; Asaah, E. Evidence that subsistence farmers have domesticated indigenous fruits (Dacryodes edulis and Irvingia gabonensis) in Cameroon and Nigeria. Agrofor. Syst. 2004, 60, 101-111, doi:10.1023/B:AGFO.0000013259.95628.22.
- Fuller, D.Q.; Asouti, E.; Purugganan, M.D. Cultivation as slow evolutionary entanglement: comparative data on rate and sequence of domestication. Vegetation History and Archaeobotany 2011, 21, 131-145, doi:10.1007/s00334-011-0329-8.
- Willcox, G. The beginnings of cereal cultivation and domestication in Southwest Asia. In A companion to the archaeology of the ancient Near East, Potts, D.T., Ed. Blackwell: Oxford, 2012; Vol. 1, pp. 163-180.
- Smith, B.D. Low-level food production. J. Arch. Res. 2001, 9, 1-43, doi:10.1023/A:1009436110049.
- Falconer, D.S.; MacKay, T.F. Introduction to quantitative genetics; Longman: Harlow, 1996; pp. 480.
- Allaby, R.G.; Fuller, D.Q.; Brown, T.A. The genetic expectations of a protracted model for the origins of domesticated crops. Proc. Natl. Acad. Sci. USA 2008, 105, 13982-13986, doi:10.1073/pnas.0803780105.
- Allaby, R.G.; Stevens, C.; Lucas, L.; Maeda, O.; Fuller, D.Q. Geographic mosaics and changing rates of cereal domestication. Philosophical Transactions of the Royal Society B Biological Sciences 2017, 372, 20160429, doi:10.1098/rstb.2016.0429.
- Allaby, R.G.; Kitchen, J.L.; Fuller, D.Q. Surprisingly low limits of selection in plant domestication. Evolutionary Bioinformatics Online 2015, 11, 41-51, doi:10.4137/EBO.S33495.
- Lewontin, R.C. The triple helix: Gene, organism, and environment; Harvard University Press: Cambridge, 2000; pp. 136.
- Cleveland, D.A.; Daniela, S.; Smith, S.E. A biological framework for understanding farmers’ plant breeding. Econ. Bot. 2000, 54, 377-394, doi:10.1007/BF02864788.
- Leach, H.M. The terminology of agricultural origins and food production systems—a horticultural perspective. Antiquity 1997, 71, 135-148, doi:10.1017/S0003598X00084623.
- Rosenswig, R.M.; VanDerwarker, A.M.; Culleton, B.J.; Kennett, D.J. Is it agriculture yet? Intensified maize-use at 1000 cal BC in the Soconusco and Mesoamerica. Journal of Anthropological Archaeology 2015, 40, 89-108, doi:10.1016/j.jaa.2015.06.002.
- Ames, O. Economic annuals and human cultures; Botanical Museum of Harvard University: Cambridge, 1939; pp. 153.
- Callicott, C.M. Interspecies communication in the Western Amazon: Music as a form of conversation between plants and people. European Journal of Ecopsychology 2013, 4, 32-43.
- Lima, A.G.M.d. A cultura da batata-doce: cultivo, parentesco e ritual entre os Krahô. Mana 2017, 23, 455-490, doi:10.1590/1678-49442017v23n2p455.
- Viveiros de Castro, E. The transformation of objects into subjects in Amerindian ontologies. Common Knowledge 2004, 10, 463-485.
- Rival, L. Seed and clone: the symbolic and social significance of bitter manioc. In Beyond the visible and the material: the Amerindianization of society in the work of Peter Rivière, Rival, L.M., Whitehead, N.L., Eds. Oxford University Press: Oxford, 2001; pp. 57-80.
- Casas, A.; Viveros, J.L.; Caballero, J. Etnobotánica mixteca: sociedad, cultura y recursos naturales en la Montaña de Guerrero; Instituto Nacional Indigenista / Consejo Nacional para la Cultura y las Artes: Ciudad de México, 1994.
- Iriarte, J.; Elliott, S.; Maezumi, S.Y.; Alves, D.; Gonda, R.; Robinson, M.; Gregorio de Souza, J.; Watling, J.; Handley, J. The origins of Amazonian landscapes: plant cultivation, domestication and the spread of food production in tropical South America. Quatern. Sci. Rev. 2020, 248, doi:10.1016/j.quascirev.2020.106582.
- Oviedo y Valdés, G.F. Historia general y natural de las Indias. [Juan Cromberger, Sevilla (1851–1855). J. A. de los Rios y Serrano (ed.)]; Imprenta de la Real Academia de la Historia: Madrid, 1535.
- Patiño, V.M. Historia y dispersión de los frutales nativos del Neotrópico; Centro Internacional de Agricultura Tropical: Cali, Colombia, 2002; pp. 655.
- Walker, J.H. Island, river, and field: landscape archaeology in the Llanos de Mojos; University of New Mexico Press: Albuquerque, 2018.
- Smith, M.E. The Aztecs; Wiley-Blackwell: Oxford, 2013; pp. 416.
- Heckenberger, M.J. Rethinking the Arawakan diaspora: hierarchy, regionality, and the Amazonian formative. In Comparative Arawakan histories: rethinking language family and culture area in Amazonia, Hill, J.D., Santos-Granero, F., Eds. University of Illinois Press: St. Louis, 2002; pp. 99-122.
- Reis, M.S.; Montagna, T.; Mattos, A.G.; Filippon, S.; Ladio, A.H.; Marques, A.d.C.; Zechini, A.A.; Peroni, N.; Mantovani, A. Domesticated landscapes in Araucaria Forests, southern Brazil: a multispecies local conservation-by-use system. Front. Ecol. Evol. 2018, 6, 011, doi:10.3389/fevo.2018.00011.
- Shepard Jr., G.H.; Clement, C.R.; Lima, H.P.; Mendes dos Santos, G.; Moraes, C.d.P.; Neves, E.G. Ancient and traditional agriculture in South America: tropical lowlands. In Encyclopedia of Agriculture and the Environment, Hazlett, R., Ed. Oxford University Press: New York, 2020; 10.1093/acrefore/9780199389414.013.597p 48.
- Fausto, C.; Neves, E.G. Was there ever a Neolithic in the Neotropics? Plant familiarisation and biodiversity in the Amazon. Antiquity 2018, 92, 1604-1618, doi:10.15184/aqy.2018.157.
- Nair, P.R. Classification of agroforestry systems. Agrofor. Syst. 1985, 3, 97-128, doi:10.1007/BF00122638.