3. Immune Alterations in Obesity and T2D
Metabolic diseases such as obesity and T2D share a common feature, augmented adiposity associated with a chronic systemic low-grade inflammation [
12,
13] which promotes the abnormal production of pro-inflammatory cytokines and the impairment of the immune response and host defense [
8]. Adipose tissue is an endocrine organ capable of secreting a diversity of cytokines and adipokines that are involved in the regulation of inflammation and homeostasis [
76]. Healthy adipocytes are sensitive to insulin, which is critical for the uptake of glucose and maintenance of blood glucose levels. In obese subjects, adipocytes increase in number and size. This phenomenon is accompanied by an insufficient vascularization of the tissue resulting in hypoxia, triggering apoptosis and/or necrosis as well as elevated secretion of more inflammatory cytokines, adipokines, and chemokines that induce a vast infiltration of immune cells contributing to lipolysis, promoting inflammation and insulin resistance. This metabolic impairment induces the initiation or aggravation of T2D [
12]. All these factors generate a low-grade inflammatory microenvironment that incites the recruitment of mast cells, neutrophils, T-cells, B-cells, and the polarization of inflammatory macrophages towards the M1 phenotype is favored in adipose tissue of obese subjects, while maintaining or even reducing the number of regulatory T cells (Treg), T helper cells type 2 (Th2), and M2 macrophages [
77]. As a result, the balance shifts from a regulatory anti-inflammatory immune state characterized for the secretion of cytokines IL-4, IL-5, IL-10, IL-13, and IL-33 to an exceedingly inflammatory state secreting multiple proinflammatory cytokines, such as IL-6, IL-8, TNF-α, IL-1β, and chemokine (C-C motif) ligand 2 (CCL2), contributing to maintain a chronic systemic inflammation [
6,
78]. Additionally, the elevated levels of IL-6 provoke an acute-phase response which increases ferritin, C-Reactive Protein (CRP) and D-dimer [
79]. The action of pro-inflammatory cytokines in immune cells can trigger signaling pathways that converge at the NF-κB and MAPK activation, resulting in the generation of intracellular inflammatory cytokine production (IL-1β, IL-6, and TNF-α) and apoptosis induction, respectively. In addition, the increased ROS production induced by excess of fatty acids and high glucose triggers the activation of NLR family pyrin domain containing 3 (NLRP3) inflammasome that, through caspasse-1 activates IL-1β form. Overall, the activation of intracellular pathways accompanied by adipose inflammation increases the production of pro-inflammatory cytokines and promotes the infiltration of more pro-inflammatory M1 macrophages. Additionally, a hallmark in obesity and T2D are the senescent immune cells which exhibit a senescence-associated secretory phenotype (SASP) characterized by secreting a huge quantity of pro-inflammatory cytokines (IL-1β, IL-6, IL-8, IL-18, CCL-2, and TNF-α) [
80,
81,
82]. SASP cells have enhanced mitochondrial apoptosis switching from apoptosis to pyroptosis, through a high mitogen-activated protein kinases (MAPKs) activity which drive IL-1 production (leading to auto-inflammation) and reduced IFN type 1 production, leading to a deficiency to combat viral infections [
83] (see ). The inflammatory cytokines also induce neuro-immune-endocrine interactions affecting the body’s response to stress. Catecholamines secreted by the sympathetic nervous system (SNS) and the adrenal glands are key regulators of metabolism and innate responses which are affected in obesity [
84,
85]. Animal models of metabolic syndrome have demonstrated the feedback mechanism between inflammatory cytokines and noradrenaline (NA) and the altered phagocytic and microbicidal capacity of macrophages, which caused a greater susceptibility to infections [
86,
87]. In fact, recent studies have proposed a novel adrenergic regulation of inflammation, via activation of β2-adrenergic receptors, highly expressed in immune cells, to modulate the inflammatory phenotype and activity profile of these cells [
85,
88].
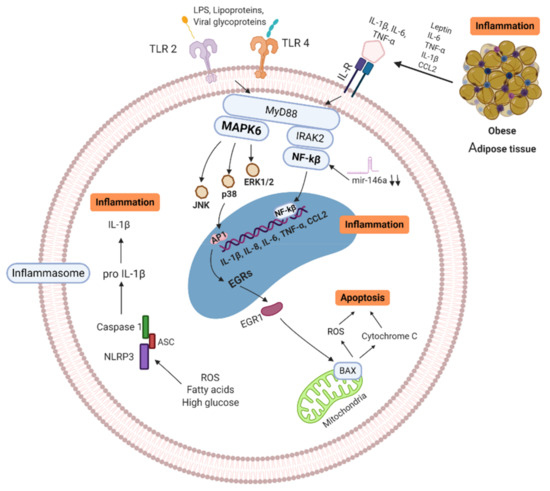
Figure 1. Signaling pathways of obese/T2D immune cell (MNC/MP). Senescence-associated secretory phenotype (SASP) cells secrete high quantity of pro-inflammatory cytokines, which trigger enhanced mitochondrial apoptosis through high mitogen-activated protein kinases (MAPKs). ROS production, free fatty acids and high glucose induce activation of NLRP3 inflammasome, activating IL-1β, leading to further inflammation. In addition, Pattern Recognition Receptors (PRR) and Interleukin Receptors (ILR) signaling through NF-κB and MAPKs pathways, resulting in more inflammatory cytokine production (IL-1β, IL-6, IL-8, IL-18, CCL-2, and TNF-α). Activation of these pathways increases the production of pro-inflammatory cytokines and promotes the infiltration of more pro-inflammatory M1 macrophages in tissues. MAPK: Mitogen-activated protein kinase, IL: Interleukins, ILR: Interleukins receptor, TNF: Tumor necrosis factor, BAX: BCL2-associated X genes, JNK: c-Jun N-terminal kinase, IRAK-2: Interleukin-1 receptor-associated kinase 2, EGRs: Early response genes, NF-κB: nuclear factor kappa B, TLR: Toll-like receptor, CCL2: C-C motif chemokine ligand 2, ROS: Reactive oxygen species, mir146a: microRNA-146a, NLRP3: NLR family pyrin domain containing 3, ASC: Adaptor protein. Created with BioRender.com.
Besides, important adipokines involved in the inflammatory process are leptin and adiponectin. Leptin is a lipostatic hormone whose main function is to regulate body weight by transmitting signals of satiety to the central nervous system and energy homeostasis [
89,
90,
91]. Evidence shows that leptin is involved in glucose metabolism, innate immune reactions and acute inflammation. The concentration of leptin in blood in obese patients with dyslipidemia is high, causing signaling alteration, promoting the development of insulin resistance and T2D [
92,
93]. In mice, it has been described an increase in leptin is associated with inflammatory markers in obese individuals [
94], and in humans production of cytokines can be induce when leptin is administered exogenously [
95]. Adiponectin is a protein whose function is to increase fat oxidation, producing a reduction in the concentration of fatty acids, increasing insulin sensitivity [
90,
96]. It is described that adiponectin levels decrease in obesity, insulin resistance, T2D and cardiovascular diseases [
97]. In addition, alteration in the relationship between leptin and adiponectin ratio has been shown to be associated with insulin resistance [
98,
99], and in the development of atherosclerosis [
100]. Consequently, leptin and adiponectin play an important role in the regulation of cardiovascular and metabolic homeostasis [
91,
96,
101].
MicroRNAs (miRNA) are other important biomarkers that play a relevant role in obesity/T2D [
102,
103,
104]. MiRNAs regulate and modulate the expression of a large number of protein-coding genes [
105]. Several studies indicate that miRNA-146a and miRNA-155 are involved in the regulation of inflammatory processes [
106,
107]. For example, miRNA-155 can promote the activation of the LPS/TNF pathway, thus contributing to the activation of the inflammatory response [
108,
109]. MiRNA-146a block the nuclear factor kappa B (NF-κB) activation induced by TNF-α and toll-like receptor ligands, therefore its function is crucial in the prevention of excessive immune response [
110]. Baldeón L et al. showed that the expression levels of miR-146a in CD14+ in T2D monocytes, was down-regulated when compared to healthy controls. The decreased expression of miR-146a in immune cells, correlates negatively with inflammatory cytokines, thus acting as a negative regulator during the activation of immune responses [
111].
Adaptive immune cells (T cells) may play a significant role in the propagation of adipose tissue inflammation in obesity. T cells from obese/T2D patients express lower levels of costimulatory molecules (CD69, CD28, CD40 ligand), and interleukin-12 receptor, as well as, produce lower levels of interferon-γ and granzyme B, compared to healthy individuals [
112,
113,
114]. It was shown in diet-induced obesity mice model that CD8+ T cells infiltrate into fat pads before macrophage infiltration. Additionally, CD8+ T cell depletion with anti CD8+ antibodies, resulted in reduced M1 macrophage infiltrations and decreased inflammatory mediators in adipose tissue, ameliorating insulin resistance and glucose tolerance [
115]. Conversely, Winer et al. found that the progression of obesity-associated metabolic abnormalities is under the pathophysiological control of CD4+ T cells. Reconstitution of CD4+ T cells, but not CD8+ T cells, in lymphocyte-free obese Rag1-null mice improved glucose tolerance, enhanced insulin sensitivity, and reduced weight gain [
116]. Furthermore, there is evidence that regulatory T cells (Treg) are key regulatory cells in adipose tissue to provide anti-inflammatory signals that block adipose tissue inflammation. Treg cells normally account for 5%–20% of the CD4+ compartment but are thought to be one of the body’s most crucial defenses against inappropriate immune responses [
117,
118,
119]. For example, Feuerer et al. demonstrated that when most of the Treg cells were ablated in fat tissue, proinflammatory transcripts were strongly expressed, suggesting the important anti-inflammatory properties of Treg in metabolic processes [
120].
B-lymphocytes have been shown to play a central role in the development of insulin resistance and glucose intolerance by activating CD4+ Th1 and Th17 cells and releasing pathogenic antibodies [
121,
122]. Epigenetic modifications such as DNA methylation play and important role in the significant increase of B cells proliferation which in turn influence the proliferation of Th17 and the production of proinflammatory cytokines [
114,
123]. For example, Ip et al. show that in vitroTh17 cells decreased upon depletion of CD19+ cells from PBMCs of T2D patients [
124]. In addition, in an animal model it was shown that the lack of mature B cells improved glucose tolerance in HFD mice [
114]. Interestingly, posttranscriptional modifications in which various glycosyltransferases are involved are well described to occur in inflammatory disease such as obesity and T2D [
125,
126]. Wu et al. described that IgG glycosylation profiles were associated with T2D, arguing that glycan score can be used as novel indicator for diabetes and inflammatory status [
127].
In conclusion, metabolic disorders such as obesity and T2D, seems to alter the balance of innate and adaptive responses, demonstrated by senescence, exaggerated cytokine inflammation, poor chemotaxis, impaired phagocytosis, glycation of circulating immunoglobulins, and abnormalities in the number and function of CD4+ T and CD8+ T lymphocytes and B lymphocytes.