Corynebacterium glutamicum is a workhorse for industrial amino acid production, and the most striking feature of this bacterium is L-glutamate secretion. L-glutamate secretion is triggered by altering cell surface structures by biotin limitation, adding surfactants, penicillin, etc. Mechanosensing by MscCG-type mechanosensitive channels is the central physiological phenomenon for the L-glutamate secretion, and understanding the structural and functional diversity of MscS bacterial mechanosensitive channel superfamily will solve long-puzzling questions in Corynebacterium glutamicum.
1. Introduction
Corynebacterial Mechanosensitive Channel Model for the Industrial L-Glutamate Production
The Corynebacterium glutamicum ATCC13032 strain was first isolated as a glutamate producer in the avian (most likely pigeon) feces-contaminated soil in 1957 by Kinoshita and Udaka [1], who described the result of an elaborated screening method with a glutamate-auxotrophic bacterium Leuconostoc mesenteroides strain, P-60 [2]. This sparked microbial amino acid production and our understanding C. glutamicum physiology has made numerous paradigm shifts in the biotechnological process. Since then, several Corynebacterium glutamicum subspecies, other than the ATCC13032 strain, have been isolated from different materials for industrial production. One of the most striking findings of C. glutamicum studies is biotin-dependent L-glutamate secretion. In nature, glutamate has two optical isomers, d(−) and l(+), and the l form (L-glutamate) is the most widely occurring, while the d form (D-glutamate) occurs rarely in the bacterial cell walls as a component of peptidoglycan layers. At a growth-limiting concentration of biotin, C. glutamicum massively releases L-glutamate into the culture media; however, this bacterium does not produce it at all under normal culture conditions [3]. Later, several alternative ways to trigger L-glutamate secretion have been reported, and the conditions are surprisingly diversified as impacting the cell wall and membrane, such as the adding of penicillin [4], ethambutol [5], ciprofloxacin [6], fatty acid ester surfactants [7], local anesthetics [8], and temperature upshift [9,10]. Correspondingly, a wide range of mechanistic models to explain L-glutamate secretion was suggested, and a coherent model was expected to elucidate the situation. Since L-glutamate is a charged amino acid, passive efflux was rejected, and an active carrier was proposed as the “carrier model” [11]. The leakage from damaged cell envelopes was considered as one of the main causes due to inhibited fatty acid biosynthesis and membrane alterations in L-glutamate production, proposed as the “leak model” [12,13,14]. Metabolism in the biotin-limited condition was shifted from energy production via tricarboxylic acid (TCA) cycle towards L-glutamate production. Thus, the “metabolic flow change model” was proposed to trigger L-glutamate secretion [15,16,17]. This puzzling situation has ended with the serendipitous discovery of the major L-glutamate exporter, NCgl1221 (cg1434). In the process of the screening of the L-glutamate overproducing strains without any specific treatment, several mutations on the NCgl1221 gene were identified as the result [18]. NCgl1221 was predicted to be a homologue of bacterial mechanosensitive channels of small conductance (MscS); however, it shows similarity only in the pore domain, and its entire structure differs from other MscS-type mechanosensitive channels [19]. This finding has emphasized the importance of bacterial mechanosensing by mechanosensitive channels on the studies of corynebacterial amino acid exporters [20,21], and mechanosensitive channels are now emergent targets for transporter engineering to use C. glutamicum as a microbial cell factory [22,23]. Conclusively, using a novel patch-clamp technique with C. glutamicum giant spheroplasts of the industrial strain ATCC13869, all endogenous mechanosensitive channels including MscCG and MscCG2 were recorded electrophysiologically in the native membrane environment [27]. These studies strongly supported the “mechanosensitive channel model” [28] and explained the L-glutamate secretion mechanisms as follows: (1) specific treatments alter membrane tension by inhibiting membrane lipids or cell wall synthesis; (2) MscCG and MscCG2 are activated by increased membrane tension;(3) L-glutamate is exported through the open pore of the MscCG-type mechanosensitive channels (). In this model, mechanosensing by MscCG-type mechanosensitive channels is the central physiological phenomenon of the L-glutamate secretion.
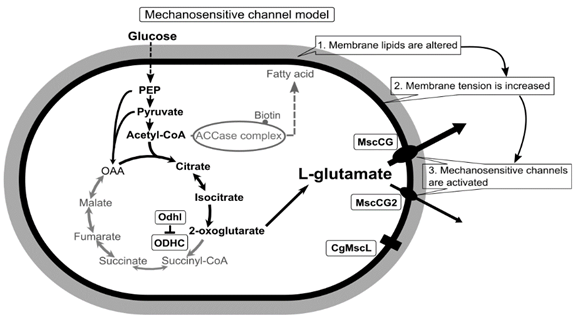
Figure 1. A scheme of the C. glutamicum L-glutamate secretion triggered by biotin limitation as mechanosensitive channel model. Biotin limitation shifts metabolic flow to produce L-glutamate by inhibiting the 2-oxoglutarate dehydrogenase complex (ODHC) activity and inhibits the acetyl CoA carboxylase (ACCase) complex activity. As a result of fatty acid biosynthesis inhibition, membrane lipids are altered to increase membrane tension. In the ATCC13032 strain, the MscS-like mechanosensitive channel MscCG is activated exclusively as major exporter by increased membrane tension to release L-glutamate. In several industrial strains, another MscS-like mechanosensitive channel, MscCG2, is also activated as a minor exporter. However, the MscL-type mechanosensitive channel CgMscL is not activated in L-glutamate secretion.
2. Diversity of MscS Mechanosensitive Channel Superfamily and the Impact of the MscCG Channel Gating on L-Glutamate Secretion
The existence of MscS-type mechanosensitive channels in the cytoplasmic membranes of both prokaryotic organisms, E. coli and C. glutamicum, indicates that these channels have developed through evolution to cope with changes of mechanical environments; however, its functions as an osmoregulator are significantly different [50]. E. coli MscS has a large open channel pore (~16 Å), and thus its conductance reaches 1 nS, whereas C. glutamicum MscCG has a significantly smaller conductance of approximately 0.3 nS [24,27,51]. As an osmoregulator, E. coli MscS has strong inactivation and adaptation mechanisms to mechanical stimuli that are needed to avoid the over-efflux of cytoplasmic molecules upon osmotic downshock [52,53]. However, C. glutamicum MscCG does not have these features and tends to be open as a metabolic valve rather than an osmotic safety valve [54]. Recent studies of the MscS channel superfamily revealed that MscS-like channels are present among cell-walled organisms, bacteria, archaea, fungi, algae, and plants, and the physiological functions of these MscS-like channels are not simply those of osmotic safety valves upon hypoosmotic shock [19]. In eukaryotes, algal and plant MscS-like channels (MSCs, MSLs) are much more complicated structures than E. coli MscS and are found in the organellar membranes of chloroplasts [55,56,57] and mitochondria [58,59,60] for mechanosensing. Fungal MscS-like channels (Msy1 and Msy2) are localized in the endoplasmic reticulum membranes and are involved in osmotic Ca2+ signalling upon hypoosmotic shock [61,62].
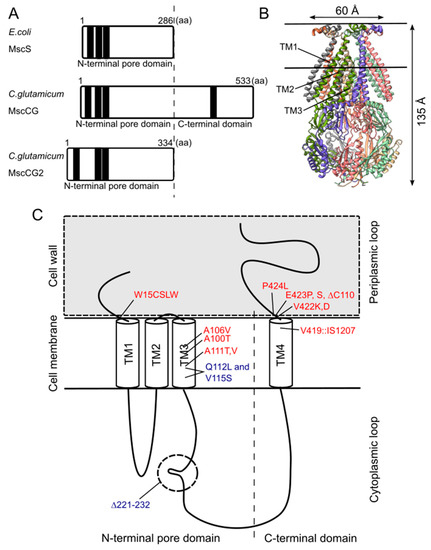
Although the physiological functions of corynebacterial MscCG-type mechanosensitive channels are still controversial, L-glutamate secretion is certainly caused by the conformational changes of the MscCG and MscCG2 channels. C. glutamicum MscCG and MscCG2 are characterized to have the N-terminal pore domain (1–286 aa), corresponding to the entire E. coli MscS that has three transmembrane helices (TM1, TM2, and TM3) and a cytoplasmic cage domain (A). Recently, Reddy et al. refined the 3D structures of the full-length E. coli MscS embedded in lipid bilayers by cryo-electron microscopy with nanodiscs [63] (B). MscCG and MscCG2 show the highest similarity in the pore-forming helix TM3 and adjacent regions. The unique feature of the mechanosensitive channel MscCG is the C-terminal domain (287–533 aa), including a cytoplasmic loop, the fourth transmembrane helix, TM4, and a periplasmic loop. In contrast, MscCG2 does not have these features (C). This structure is highly conserved only in corynebacteria. Several gain- and loss-of-function mutations on MscCG were identified by the glutamate productivity assay and bacterial patch-clamp technique (C). Originally, Nakamura et al. identified W15CSLW, A100T, A111T, A111V in the N-terminal pore domain and V419::IS1207, P424L in the periplasmic loop during the screening of L-glutamate overproducing strains. These mutations were reported to cause a spontaneous L-glutamate secretion in C. glutamicum [18]. Afterwards, Nakayama et al. reported using E. coli patch-clamp that the spontaneous L-glutamate secretion was caused by the gain-of-function mutation on mechanosensitivity of MscCG channels [64]. These findings proved that MscCG mechanosensitivity can be evaluated by L-glutamate productivity in C. glutamicum. Using these assays, the functional domain of MscCG has been thoroughly investigated. Yamashita et al. reported, using L-glutamate productivity assay, that the N-terminal pore domain (1–286 aa) is essential to export L-glutamate, and suggested, using homology modelling, that MscCG has an extra small loop structure (221–232 aa) and its deletion resulted in the loss of channel functionality [65]. Becker et al. reported additional point mutations in the N-terminal pore domain and the impact of the C-terminal domain on the channel function [66,67]. A106V was the gain-of-function mutation to cause the spontaneous L-glutamate secretion, and interestingly, Q112L and V115S double point mutation caused a loss-of-function mutation, such that C. glutamicum cannot export L-glutamate, even with penicillin treatment. In contrast, the deletion of the periplasmic loop (423–533 aa) in the C-terminal domain caused spontaneous L-glutamate secretion, but not the further deletion of the periplasmic loop and the fourth transmembrane helix TM4, indicating that the fourth transmembrane helix TM4 is involved in the mechanosensitivity of MscCG channels. Moreover, Krumbach et al. further identified the gain-of-function mutations V422K, V422D, E423P, and E423S using CRSPER/Cas12a genome-editing technology [68]. Based on the position of these mutations, the interaction with other components of the cell wall was suggested to be crucial for MscCG mechanosensitivity.
Figure 2. Structural features of the mechanosensitive channel MscCG. (A). Secondary structure comparison among E. coli MscS, C. glutamicum MscCG, and MscCG2. Black bars show predicted transmembrane (TM) helices by TOPCONS program (https://topcons.net/). (B). The cryoEM 3D structure of E. coli MscS embedded in nanodiscs with POPC:POPG = 1:4 (structure was cited from the Protein data bank 6PWP). (C). The domain structure of the MscCG channel and the gain (red)-and loss(blue)-of-function mutations.
This entry is adapted from the peer-reviewed paper 10.3390/microorganisms9010201