Mycoplasma pneumoniae is a major causative agent of community-acquired pneumonia which can lead to both acute upper and lower respiratory tract inflammation, and extrapulmonary syndromes.
- Mycoplasma pneumonia
- virulence factors
- pathogenesis
- adhesins
1. Introduction
Community-acquired pneumonia (CAP) is associated with high morbidity and mortality, and the disease is also a major threat to public health worldwide [1]. About 8–40% of CAP in children admitted to hospitals were caused by Mycoplasma pneumoniae [2,3,4]. Based on the reported cases in China, M. pneumoniae infections accounted for 19.2% of all CAP cases in adults, and the prevalence of CAP in children and teenagers, ranged from 10% to 30% [1,5]. In the USA, a recent study of 2254 hospitalized children with CAP showed that 8% children with median age of 7 years were positive for M. pneumoniae by polymerase chain reaction (PCR) [6].
Airborne droplets containing M. pneumoniae can be transmitted and spread among people through coughing and sneezing. M. pneumoniae causes both upper and lower respiratory tract infections, and in most cases the clinical symptoms are non-specific [7]. Tracheobronchitis is the most common type of lower respiratory infection, the incidence of which is about 20 times that of pneumonia, and 10–40% of respiratory tract infections caused by M. pneumoniae will eventually develop into pneumonia [8]. While most pneumonia caused by M. pneumoniae (MPP) cases are benign, some cases may develop into severe pneumonia and refractory pneumonia with pleural effusion, multi-organ dysfunction, and serious long-term sequelae, including bronchiolitis obliterans and bronchiectasis [9]. Although CAP is the most significant disease caused by M. pneumoniae, the pathogen is known to cause upper respiratory tract infections. Pharyngitis is commonly reported while rhinosinusitis and otitis media are less frequently encountered in upper respiratory tract infections caused by M. pneumoniae [7].
M. pneumoniae respiratory infections are associated with asthma exacerbation during which patients will suffer from a combination of symptoms including sudden or progressive coughing, respiratory distress, wheezing or chest pain [10,11]. The onset of asthma is due to the release of Mycoplasma-mediated cytokine in infected patients [12]. Respiratory infections caused by M. pneumoniae are also associated with a wide array of extrapulmonary manifestations such as meningoencephalitis, myocarditis, nephritis, atherosclerosis and mucocutaneous eruptions, etc. [13,14,15,16,17]. More importantly, M. pneumoniae induces mucocutaneous diseases include Stevens-Johnson syndrome and M. pneumoniae-associated mucositis. These mucocutaneous diseases are frequently associated with systemic inflammation and higher risk of the occurrence of long-term sequelae [18,19,20,21].
Due to the atypical symptoms produced during M. pneumoniae infection, pneumonia can be underestimated during the early stage of infection. There are no distinctive clinical or radiographic features in patients with M. pneumoniae infections, so laboratory diagnosis mainly based on rapid culture of throat swab specimens, PCR and serological assays. Furthermore, enzyme-linked immunosorbent assays (ELISA) detecting the N-terminal fragment of P116 protein and the C-terminal region of P1 protein both hold promise for serodiagnosis [22,23]. The IgM ELISA assays based on the short recombinant P116 and P1 proteins were shown to improve the specificity of the immunodiagnostic assay [22].
Although M. pneumoniae infection is generally self-limiting and does not require antibiotic treatment, patients of all age groups can develop severe, life-threatening or extrapulmonary diseases [24]. Antibiotics such as tetracycline and fluoroquinolone have been reported to be effective in eliminating M. pneumoniae infections [25] but tetracyclines cause discoloration of bones and teeth in young children. Fluoroquinolones can also affect the muscle, joint and tendon. Instead, macrolides, which have fewer side effects, have been the drug of choice for treating M. pneumoniae infection in past years [26]. More worrisome is that the extensive use of macrolides in China has led to a particularly high rate of macrolide resistance in this organism (69%~95%) [27]. The emergence of antibiotic resistance represents another challenge regarding the treatment of M. pneumoniae infections. Failure in antibiotic treatment has caused an increase in mortality rate during recent years [28]. Although the clinical outcomes of infections caused by macrolide-susceptible and -resistant M. pneumoniae isolates are not significantly different, patients infected with macrolide-resistant isolates had a longer febrile period (1.71 days), length of hospital stay (1.61 day), antibiotic drug courses (2.93 days), and defervescence time after macrolide treatment (2.04 days) compared to patients infected with macrolide-sensitive isolates [29]. Furthermore, macrolide-resistant strains may be associated with more extrapulmonary complications, and severe clinical and radiological features [24,30]. Hence, the development of vaccines against M. pneumoniae infections is a potential solution for the prevention of infections caused by the pathogen.
2. Virulence and Pathogenesis of M. pneumoniae
M. pneumoniae encodes a variety of virulence factors, which include adhesins, glycolipids, toxic metabolites, community-acquired respiratory distress syndrome (CARDS) toxin, and capsular polysaccharides. Table 1 summarizes the key virulence factors associated with M. pneumoniae.
Table 1. Key virulence factors of M. pneumoniae.
| Pathogenic Mechanism | Virulence Factor | Gene Annotation | Reference |
|---|---|---|---|
| Adherence | P1 | MPN141 | [31] |
| P30 | MPN453 | [32,33] | |
| P40 (Protein C) | MPN142 | [34] | |
| P90 (Protein B) | MPN142 | [34] | |
| P200 | MPN567 | [35] | |
| Hypothetical protein HMW1-3 (high molecular weight) | MPN447/310/452 | [36] | |
| P116 | MPN213 | [36] | |
| P65 | MPN309 | [37] | |
| Elongation factor thermo unstable (EF-Tu) | MPN665 | [38,39,40] | |
| Pyruvate dehydrogenase subunit B | MPN392 | [41] | |
| Glycolytic enzymes enolase | MPN606 | [41,42] | |
| TopJ | MPN119 | [43] | |
| Immune evasion | Nuclease | MPN491 | [44] |
| Immunoglobin binding protein (IbpM) | MPN400 | [45] | |
| Inflammation injury | H2O2 | / | [46] |
| Reactive oxygen species (ROS) | / | [46] | |
| H2S | / | [47] | |
| HapE enzyme | MPN487 | [47,48] | |
| Oxidase GlpO | MPN051 | [49] | |
| Membrane lipids | / | [50] | |
| Membrane lipoproteins | / | [51] | |
| Capsular materials | / | [52] | |
| Cytotoxicity | Community-Acquired Respiratory Distress Syndrome (CARDS) toxin | MPN372 | [53,54] |
| Cytotoxic nuclease | MPN133 | [55] | |
| Gliding motility | P65 | MPN309 | [37] |
| P30 | MPN453 | [32] | |
| Hypothetical protein MPN387 | MPN387 | [56] | |
| P24 | MPN312 | [57] | |
| P41 | MPN311 | [57] |
2.1. Adhesins
M. pneumoniae attaches to epithelial cell surfaces with a high affinity for human respiratory epithelial cells. The pathogen has no cell wall and colonizes the respiratory tract via its specific attachment organelle, which is a protrusion at one end of the Mycoplasma pneumoniae cell (Figure 1). The attachment organelle consists of internal and surface structures [58]. The internal structure is made up of a dumbbell-shaped terminal button consisting of three protein molecules (HMW2, HMW3, and P65), paired plates (HMW1, HMW2, CpsG, and HMW3), and a bowl complex (Lon, P24, TopJ, P200, P41, MPN387, and HMW2). The Nap structure in the surface adhesion complex consists of the main adhesins (P1 and P30) and accessory proteins (P40 and P90) surrounding the cell membrane (Figure 1). During gliding, the force generated at the bowl complexes is transmitted through the paired plates and reaches the P1 adhesin complex [58]. P30 adhesin is a membrane protein at the distal end of the attachment organelle, required for cytoadherence, gliding motility and stabilization of the accessory protein P65 [33]. Interaction of the M. pneumoniae attachment organelle with the host’s respiratory epithelium induces cytoskeleton rearrangement in the host cell, which promotes intracellular delivery of the pathogen [59,60].
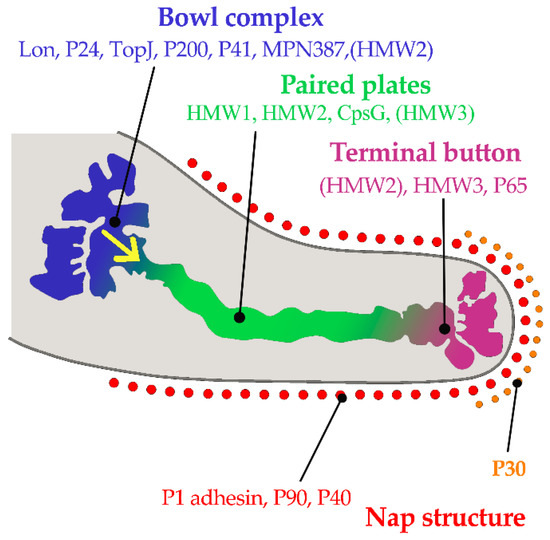
Figure 1. Component proteins of the internal structure of attachment organelle and proposed mechanism of movements for gliding in M. pneumoniae. HMW1, HMW2, and HMW3 refer to three high molecular weight (HMW) proteins. The force is generated at the bowl complexes, transmitted through the paired plates, and reaches the P1 adhesin complex in the direction of the yellow arrow. (Based on ideas from Nakane, et al. [58]). Copyright: ©2015. Public Library of Science. Creative Commons Attribution License and disclaimer available from: http://creativecommons.org/licenses/by/4.0/.
The host receptors for M. pneumoniae are sialylated glycoproteins on the respiratory epithelium. The nature and density of host receptor moieties affect the attachment and gliding mobility of the pathogen. P1 adhesin binds to both α-2,3 and α-2,6 linkages, but only the latter type of linkage supports gliding of M. pneumoniae [61].
Attachment and invasion of M. pneumoniae produces direct damage to the host’s respiratory epithelium [59,62]. Disturbance of carbohydrate metabolism, amino acid intake and protein synthesis of the host cell results in nutrient depletion [46] Furthermore, the oxygen radicals generated by the pathogen in the host cell can lead to cilia destruction and host cell damage [63,64].
2.2. Inflammation Injury
Bacterial cellular components, metabolites and toxins released from M. pneumoniae are able to induce damage in the host tissues. These include cytotoxicity, oxidative damage, apoptosis and immune-pathological damage.
2.2.1. Enzymes and Metabolites
The enzyme, HapE, of M. pneumoniae is a virulence factor that can produce H2S by the desulfurization of cysteine [47,48] which can lead to erythrocyte lysis. This enzyme mediates inflammatory reactions via adenosine triphosphate (ATP)-sensitive K+ channels [65]. Oxidation of glycerol by the pathogen produces toxic metabolites [66] including hydrogen peroxide [67,68] which injures cells by causing inflammation. In addition, the Ca2+-dependent cytotoxic nuclease (encoded by MPN133) produced by M. pneumoniae can lead to apoptotic-like programmed cell death in the host.
2.2.2. Lipoproteins
More than 50 different lipoproteins have been identified in M. pneumoniae, many of them involved in inflammatory reactions [69]. The transcription of M. pneumoniae lipoprotein genes are regulated in response to changes in environmental conditions (e.g., oxidative and acidic stress) [70,71]. The N-terminal region of all the lipoproteins contains a lipid-cysteine structure and these lipoproteins induce inflammation [69]. M. pneumoniae lipoproteins can be recognized by toll-like receptor (TLR)1, TLR2 and TLR6, which stimulate the release of proinflammatory cytokines including tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6 and other inflammatory mediators via the nuclear factor κB (NF-κB) pathway [72,73].
2.2.3. Community-Acquired Respiratory Distress Syndrome (CARDS) Toxin
The CARDS toxin encoded by MPN372 is a unique bacterial adenosine diphosphate (ADP)-ribosylating and vacuolating toxin produced by M. pneumoniae [74,75]. The structure of CARDS toxin comprises a triangular molecule in which N-terminal mono-ADP ribosyl-transferase (mART) and C-terminal tandem β-trefoil domains associate to form a unique overall architecture different from other well-recognized ADP-ribosylating bacterial toxins [53]. CARDS toxin demonstrates high binding affinity to human surfactant protein A and annexin A2 when present in the airway epithelia and exhibits specific biological activities including mono-ADP ribosylation and vacuolization [53,74]. CARDS toxin binds to mammalian cell surface receptors and is internalized rapidly in a dose and time-dependent manner. The internalization process is mediated by clathrin molecules, which form a molecular scaffold for uptake of CARDS toxin [76]. The toxin is cytotoxic to mammalian cells by activation of the NLRP3-associated inflammasome and further promotes the release of IL-1β and IL-18 [77,78,79]. CARDS toxin increases the expression of the proinflammatory cytokines IL-1β, IL-6 and TNF-α in a dose- and activity-dependent manner [80]. CARDS toxin is capable of inducing an allergic-type inflammation in animals [81,82], but there is no convincing evidence that CARDS toxin is a causal factor of M. pneumoniae-associated asthma.
2.2.4. Lipids
The cell membrane of M. pneumoniae has a high lipid content (comprising primarily of the acidic glycerophospholipids phospholipids and cholesterol), which can infiltrate the host epithelial cells, disrupt the lipid bilayer of the cell membrane and cause leakage of ionic metabolites [54,55]. Furthermore, some scholars speculate that these lipids may act as potential TLR4 ligands for binding to TLR4 and elicit macrophage autophagy, eventually leading to the secretion of proinflammatory cytokines [50,83] and triggering typical host cell inflammatory responses [50,84].
2.2.5. Capsules
M. pneumoniae has a capsular structure made up of polysaccharides [52] which may be potential virulence factors and are immunogenic, but its functional role in pathogenesis remains unclear and needs to be further explored [85,86,87].
2.3. Immune Evasion
M. pneumoniae has multiple strategies to escape host immune responses in order to ensure survival of the pathogen. Its survival includes immune evasion which may play an important role in pathogenesis. Inadequate immune responses against the invading pathogen results in uncontrolled proliferation and host tissue damage [88].
2.3.1. Molecular Mimicry
The term molecular mimicry can be described simply as “pathogens sharing a structural relationship with the host are tolerated as self, just like constituents of the host” [2,88,89]. The immune response targets the pathogen-peptide mimicking the host’s self-antigen, leading to the activation of naive, autoreactive T-cells specific to the corresponding self-antigen [89]. M. pneumoniae antigen mimics host cell components, thus the host immune response induced by the pathogen causes auto-immune responses and injuries to multiple organs [2,90].
The C-terminal region of the P1 and P30 proteins in M. pneumoniae show high levels of homology to troponin, cytoskeletal proteins, keratin and fibrinogen of the host [46,91]. Antibodies produced in response to M. pneumoniae infections will target various host tissues and form immune complexes, which aggravates the autoimmune response, leading to inflammatory injuries in the extrapulmonary tissues [13,46].
2.3.2. IbpM
Immunoglobin binding protein (IbpM) is a surface protein encoded by MPN400 that binds strongly to various immunoglobulins (IgM, IgG, and IgA) produced by the host [45]. Blötz et al. demonstrated that IbpM was required by M. pneumoniae to produce cytotoxic effects in host cells and is thus regarded as a virulent factor [45].
2.3.3. Antigen Variation
It has been observed that the surface adhesins P1, P40, and P90 of Mycoplasma pneumoniae display sequence variation [92,93]. Sluijter et al. demonstrated that the RecA protein homolog encoded by MPN490 promoted gene exchange between homologous DNA sequences (RepMP) in M. pneumoniae [94]. The RepMP are repetitive sequences present within genes encoding surface proteins such as the adhesins. Homologous recombination between these RepMP sequences generates sequence changes within the adhesin genes, which results in variations of surface adhesins and facilitates evasion of host immune surveillance [94,95,96].
The role of post-translational modifications of M. pneumoniae-specific proteins (e.g., P1, P40, P90) is a relatively new aspect of bacterial epigenetics [34]. The posttranslational modification of cytoadherence proteins by the protein kinase PrkC is essential for the development and function of the M. pneumoniae terminal organelle [97]. P1 adhesin of M. pneumoniae M129 is subject to extensive post-translational processing forming 22 proteo-forms, which are specific molecular forms of a protein product arising from a specific gene. Each of the proteo-forms retain the ability to bind to host molecules or their structural mimics and are surface accessible [31]. There are many issues that require further study, such as whether the antigen variations caused by post-translational modifications can affect the pathogenicity of M. pneumoniae.
2.3.4. Intracellular Survival
M. pneumoniae can survive for a long time in the human lung carcinoma cell (A549) [98], but the pathways related to intracellular survival remain to be elucidated. Intracellular M. pneumoniae has mechanisms to protect the pathogen against phagocytosis and antibiotics. This may explain why M. pneumoniae infection can develop into chronic lung disease, such as refractory pneumonia caused by macrolide-resistant M. pneumoniae due to the lack of timely and effective antibiotic treatment.
2.3.5. Others
Moreover, M. pneumoniae has an antioxidant mechanism to protect against oxidative reactions such as reactive oxygen species (ROS) damage [46,99]; A nuclease encoded by MPN491 can degrade neutrophil extracellular traps (NETs), which helps the pathogen to escape from the immune attack of host cells [44].
In summary, the pathogenesis of M. pneumoniae involves mainly the following four factors: immune evasion, adhesion, inflammatory injury and cytotoxicity. Figure 2 shows these four key pathogenic mechanisms of M. pneumoniae infection.
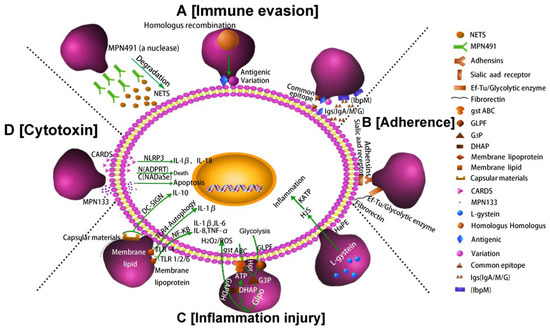
Figure 2. Pathogenic mechanisms of M. pneumoniae. (A) Nuclease and IbpM in M. pneumoniae enable immune evasion, and homologous DNA recombination leads to antigen variation; (B) M. pneumoniae adhesion causes cell damage. Additionally, the P1 adhesin protein binds to the sialic acid receptor on the host cell surface contributing to M. pneumoniae adherence and gliding. Furthermore, elongation factor Tu (EF-Tu) can bind strongly to a diverse range of host molecules (such as fibronectin), contributing to adhesion; (C) Inflammation-inducing factors (HapE enzyme, oxidase GlpO, membrane lipids, lipoproteins, and capsular materials) activate host cell inflammatory pathways; (D) M. pneumoniae secretes cytotoxic nuclease (catalytic protein encoded by MPN133) and CARDS toxin.
This entry is adapted from the peer-reviewed paper 10.3390/pathogens10020119