Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Zoology
Trichuriasis is the clinical disease of animals infected with the parasite of the genus Trichuris. This review attempts to present information on Trichuris spp. infestation in neo-tropical rodents that are utilized for meat consumption by humans
- agouti
- lappe
1. Introduction
The neo-tropics is a geographical region located in the western hemisphere between the Tropic of Cancer and the Tropic of Capricorn. Geographical territories present within this zone include the southern parts of North America, all of Central America, the northern parts of South America, and all of the Caribbean [1]. Animals that are present in this region can be categorized into three groups: imported domesticated animals [2], domesticated animals originating from the neo-tropics [3], and non-domesticated neo-tropical animals [4]. For the purpose of this review, neo-tropical rodents that are included belong to the domesticated and non-domesticated groups. Domesticated neo-tropical rodents, such as the guinea pig, are utilized in South America for their meat and are reared in captivity to provide meat protein for rural villages. The guinea pig is able to utilize household waste and provide income and food for these communities [5,6]. Neo-tropical rodents on the verge of domestication are the agouti, lappe, and capybara. These animals have been reared in captivity in South America and the Caribbean for their meat [1]. These animals have been able to breed in captivity: the agouti produces four offspring per year [7], the lappe produces two offspring per year [8], and the capybara can produce eight offspring per year [9,10]. These animals are ideal in that they can utilize local feed resources and are adapted to local conditions of high heat and humidity. The meats produced by these rodents are highly nutritious, with high protein values and low fat and cholesterol concentration [11,12,13,14]. Trichuris spp., also known as whipworms, have parasitized many domesticated species, causing enteritis, diarrhea, and weight loss [15]. Trichuris spp. adults live in the caecum and colon; this predilection site has occurred due to evolution. The life cycle is direct; eggs with characteristic bi-polar plugs are passed in the feces and take two to three weeks to become infective (Figure 1) [16]. Animals become infected by the ingestion of infective eggs [16]. However, there has been limited information on the effects of Trichuris spp. on neotropical rodents (domestic and semi-domestic). Thus, the objective of this review is to summarize the species of Trichuris that parasitizes these rodents, the effect of this parasite on these animals, and the zoonotic potential of this pathogen.
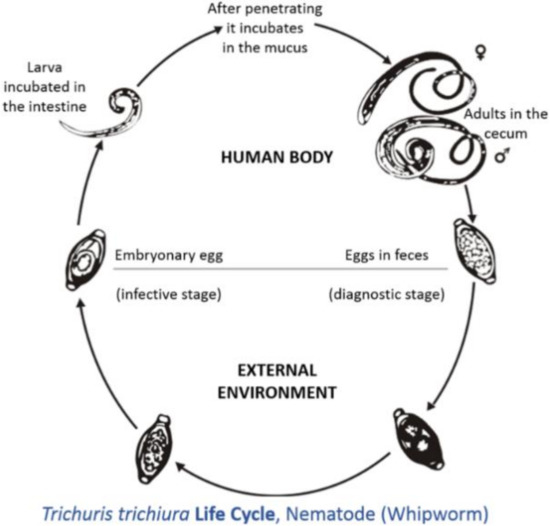
Figure 1. Life cycle of Tichuris trichiura (taken from [17]).
2. Trichuris spp. of Veterinary and Public Health Importance
2.1. Trichuriasis of Man
Trichuriasis is one of the major infectious diseases of children in developing countries [18]. Trichuris trichiura is a major, soil-transmitted helminth targeted by the World Health Organization in their mass drug administration program for pre-school and primary school children in endemic developing countries [18]. There have been several cases of trichuriasis reported in humans. In some cases, it has been due to three Trichuris spp.: T. trichiura, T. vulpis, and T. suis. Humans have been infected with T. vulpis, and the diagnosis was made based on the morphology of the eggs and vulva from an adult female [19]. Molecular techniques were used on Trichuris spp. egg present in feces to identify T. suis and T. trichiura in human populations from Thailand [20]. T. suis has been experimentally given to humans, and the author stated that feces were negative for Trichuris eggs 40 days post-infection [21]. Experimentally treated patients showed no symptoms of gastrointestinal distress [21]. In contrast to the previous studies, Kradin et al. [22] showed that iatrogenic infection with T. suis resulted in a persistent active infection in man. Pathological findings from colonic biopsies showed several round helminths beneath the ileocecal mucosa epithelium [22].
Trichuris trichiura has human and non-human primates as its natural hosts [23]. Mixed infections with various Trichuris spp. in humans have been documented. There have been cases of mixed infections with T. vulpis and T. trichiura [24,25]. The identification of the species of Trichuris spp. was based on the morphology of eggs [24] and polymerase chain reactions of the helminth eggs [25]. Trichuris trichiura and T. vulpis was also found in the stool samples of dogs that roamed around the community. This shows that dogs are key to the transmission of Trichuris spp. to humans, but further work needs to be done to validate this finding [25].
Infections with T. vulpis have been reported in children and adults [19,26,27]. However, all cases of trichuriasis in humans caused by T. vulpis have had some association with dogs, and the diagnosis was made based on morphology of eggs present in the feces. Clinical signs reported in humans are abdominal discomfort, epigastric pain, nausea, vomiting, diarrhea, and poor appetite [24]. Patients with T. vulpis [24,26,27] and T. trichiura [19] have been treated with mebendazole and albendazole with improvements of clinical signs [19,24,26,27]. However, in vivo studies on albendazole and mebendazole have shown little efficacy against T. trichiura [28]. At 14 days post-treatment, there was no difference in the disease prevalence seen between treatments of patients with 400 grams of albendazole [28]. Therefore, alternative anthelmintic treatment against T. trichiura should be investigated. Ivermectin has been used to treat Trichuris spp.; however, it is very ineffective, as these parasites have become resistant to this drug. However, due to the increased prevalence of anthelmintic resistance, the drugs used to treat trichuriasis should be done with caution.
2.2. Morphological and Molecular Identifications of Trichuris spp.
2.2.1. Morphological Identification of Trichuris spp. in Pigs, Dogs, Cats, Humans, and Non-Human Primates
Morphological analysis of Trichuris spp. has been used for identification within various host species. Trichuris trichiura infection has been investigated in humans, non-human primates, and pigs, but based on morphological analysis, the T. trichiura found in humans and non-human primates were indistinguishable [29]. In pigs, T. suis was differentiated from T. trichiura, based on the lack of peri-cloacal papillae in adult specimens. In female specimens, there were no morphological differentiation between T. suis and T. trichiura [29]. Ruminants evaluated in India using morphological analysis identified T. ovis as the major parasite [30].
Further research was done in domestic cats in St. Kitts. Based on the size of the Trichuris spp. identified, authors believed that it was T. campanula, but based on the vulva structure the authors confirmed it was T. serrata. In conclusion, the authors, identified the parasite as T. serrata, but recommended that molecular studies must be done in order to reliably identify this parasite [31]. In dogs, male and female adult T. vulpis could be identified based on nine parameters (including body length, length of cuticular processes, and width of body at tail part) [32]. Male T. vulpis can be distinguished from other species by spicule sheath ornamentation (the dimensions of the spicule) [32].
Recently, the morphometric approach analyzing the adult worms and eggs of Trichuris spp. of non-human primates were analyzed [33,34]. Morphometric data on the adult worms showed that features present in the females made them indistinguishable for species characteristics, but adult male worms may be used to differentiate Trichuris populations [33]. Geometric morphometric analysis is a new diagnostic tool that can be used to differential Trichuris spp. present in non-human primates. However, further data must be collected to determine the sensitivity and specificity of this diagnostic tool [34]. Combination of various techniques, such as the use of molecular and morphological analysis, should be performed for confirmation of various Trichuris spp. [33].
2.2.2. Molecular Identification of Trichuris spp. in Domestic and Non-Domestic Ruminants
Molecular techniques have been used to identify various Trichuris spp. in their animals or human hosts. Such techniques have been applied to Trichuris spp. found in ruminants (both domesticated and non-domesticated). Four Trichuris spp.—T. discolor, T. ovis, T. globulosa and T. skrjabini—have been identified as inhabiting the caecum and colon of ruminants [35,36,37,38,39,40,41,42,43,44,45]. One of the major discoveries was the identification of T. globulosa and T. ovis as the same species by isoenzymes [35], using second, internally transcribed spacer ribosomal DNA (ITS2 rDNA) [38] and ITS1-5.8S-1TS2 [37]. Further molecular analysis was done comparing T. ovis and T. discolor, where the entire mitochondrial DNA (mtDNA) was analyzed [42], and with the use of internally transcribed spacers 1, 2, and 16S, partial DNA sequencing (ITS1, 2, 16rDNA) was completed [44]. Based on mtDNA and rDNA, T. ovis and T. discolor can be classified as two different species.
Trichuris skrjabini, found in small ruminants (sheep and goats), was characterized using isoenzymes [36], ITS1-5.8S-1TS2 [37], and cytochrome oxidase subunit 1 and mitochondrial 16S rDNA [39]. Authors have stated that T. skrjabini is an independent species but has close relations to other Trichuris spp. that parasitize small ruminants. Trichuris discolor has been identified in domestic ruminants with the use of molecular techniques; however, it was recently identified in wild ruminants, such as the roe deer (Capreolus capreolus), sika deer, (Cervus nippon), red deer (Cervus elephus), fallow deer (Dama dama), and mouflons (Ovis orientalis musimon) [43,44,45]. In wild ruminants, T. discolor was identified with use of ITS1-5.8S-1TS2 [43,44,45], but in cattle different populations of T. discolor in Iran, Spain, and Japan were investigated using 16S partial gene mtDNA, as well as ITS1 and 2 [43]. Callejon et al. [43] noted that there were specific populations of T. discolor groups based on geographical location. The author noted that one reason may be due to two cryptic species of T. discolor from Japan and Iran, as well as another from Spain.
2.2.3. Molecular Identification of Trichuris spp. in Cats, Dogs, Pigs, Humans, and Non-Human Primates
Trichuris spp. has also been identified molecularly in pets, such as dogs and cats. In cats it is associated with typhlitis, which also occurs in other animals [46]. Identification of T. serrata (cats) and Trichuris vulpis (dogs) was accomplished through the use of 18S rDNA (cats) and enzyme-linked immunosorbent assay (ELISA) and ITS1-5.8S-1TS2 (dogs) [47,48,49]. Comparative genetic studies were done of the T. vulpis found in dogs and T. suis found in pigs (wild and domesticated). There was a difference seen in amplified ITS1-5.8S-1TS2 rDNA between the T. vulpis found in dogs and T. suis found in pigs. Interestingly, T. suis collected from wild pigs (Sus scrofa scrofa) and domestic pigs (Sus scrofa domestica) showed no sequential genetic differences [49].
Several non-morphological processes were used to identify T. suis found in pigs using isoenzymes [50], ITS 1 and ITS2 regions of rDNA [51], large mitochondrial subunits and ITS2 [52], and nuclear ribosomes (18S, ITS2) [18]. Due to the zoonotic potential of T. suis and its morphological similarity to T. trichiura previous molecular studies have been done in both human and non-human primates [53,54,55]. Trichuris spp. was taken from pigs (wild and domestic) and non-human primates (Colobus guereza kikuyensis and Nomascus gabriellae) and analyzed by amplification of rDNA (ITS1-5.8S-1TS2). The authors confirmed that the T. suis found in pigs was genetically different from T. trichiura in Colobus guereza kikuyensis and Nomascus gabriellae [53]. Nissen et al. [54] conducted a similar study to Cutillas et al. [53], but T. suis and T. trichiura were identified in pigs and humans in Uganda. The gastrointestinal tract of pigs only contained T. suis, while in humans T. trichiura, T. suis, and a heterozygous type was identified [54]. This showed that the use of ITS 2 and β-tubulin allowed the identity of several species of Trichuris in humans to be highlighted.
The research done by Cutillas et al. [53] and Nissen et al. [54] highlights the fact that humans and non-human primates may be infected with several species of Trichuris that are generally classified as T. trichiura. This was seen with Trichuris spp. samples taken from the wild Japanese macaques (Macaca fuscata), where the Trichuris spp. identified had genetic (18S rDNA) dissimilarity compared to those found in humans [56]. This new hypothesis sparked scientists to investigate this phenomenon at a molecular level (Figure 2). Ravasi et al. [57] investigated the genotype of human and non-human primates in Central Africa. Sequencing of the rDNA (ITS1-5.8S-1TS2) revealed two Trichuris genotypes that infect both humans and non-primates [57]. Ghai et al. [58] found similar results to Ravasi et al. [57], but three Trichuris genotypes were identified as circulating within human and non-human primates. Humans were infected with two genotypes: one genotype that was only common to human samples (Group 1), and another genotype that infected humans as well as non-human primates (black-and-white colobus (Colobus guereza), blue monkeys (Cercopithecus mitis), grey-cheeked mangabeys (Lophocebus albigena), l’hoest monkeys (Cercopithecus lhoesti), olive baboons (Papio anubis), red colobus (Procolobus rufomitratus), red-tailed guenons (Cercopithecus ascanius), and the chimpanzee (Pan troglodytes)) (Group 3). The intermediary group (Group 2) had a Trichuris genotype that affected non-human primates (black-and-white colobus (Colobus guereza and the red colobus (Procolobus rufomitratus) [58]. Furthermore, this new species of Trichuris was found in the Francois’ leaf monkey (Presbytis francoisi) and the Colobus guereza kikuyensis using mtDNA, rDNA, and morphometry [59,60].
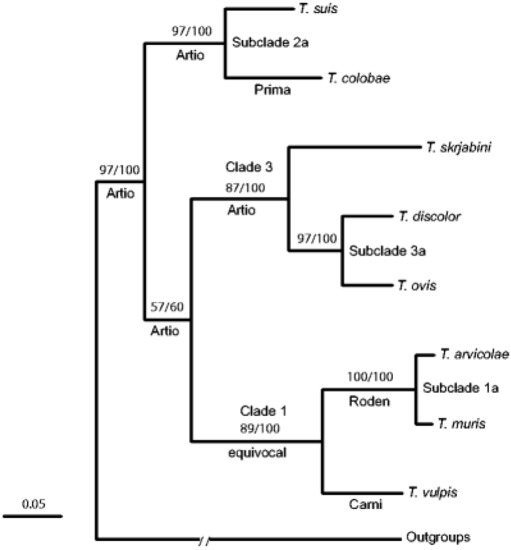
Figure 2. Phylogenic tree of Trichuris spp. (taken from Cutillas et al. [53]).
2.2.4. Molecular Identification Trichuris spp. in Rodents
Trichuris spp. has been found in domestic livestock and pets, but there are also species that are specific to rodents. The initial molecular research that was done on the Trichuris spp. present in rodents focused on European rodents [61]. Trichuris muris was identified in Murid rodents in Europe with the use of rDNA (ITS1-5.8S-ITS2). It was found that two lineages had occurred, due to geographical distribution. One was found in northern Spain to Denmark, and the other in the Southern Europe (Croatia, Romania, and Turkey) [61]. In recent years, several new species of Trichuris arvicolae have been found in Arvicolinae rodents using multi-local enzyme electrophoresis [62] and rDNA (ITS1-5.8S-ITS2) [63]. Further investigations were done in the phylogeographic analysis of T. arvicolae in Europe, using the mtDNA cytochrome subunit 1 gene (cox1) and rDNA (ITS1-5.8S-ITS2). Nuclear genetics (ITS1-5.8S-ITS2) suggest that T. arvicolae show two geographic and genetic lineages (Neoarctic and Palaearctic). Mitochondrial results gave further details into the Palaearctic region, giving three geographic and genetic lineages (Northern Europe, Southern and Eastern Europe, and Italy and France) [64].
Scientists also investigated Trichuris present in Sigmodontinae rodents in South America (Argentina). New species, such as Trichuris novonae, were identified based on morphological analysis [65]. Another species that was identified morphologically was T. pardinasi [64]. Based on molecular characteristics, using ITS2 (rDNA), a new species named Trichuris bainae was identified [66]. Molecular analysis using cox1 and mitochondrial cytochrome b (cob) on the Trichuris spp. found in Sigmodontinae rodents found three clades corresponding to three different species, which were T. pardinasi, T. bainae, and T. navonae) [67]. Further to this, T. massoiai was identified in Holochilus chacarius (Cricetidae: Sigmodontinae) using morphological mitochondrial (cox1 and cob) and nuclear (ITS2) markers [68].
Callejon et al. [41,69] investigated nuclear (18S, triose phosphate isomerase) and mitochondrial (cox1, cob1) genes from Trichuris spp. from nine various host species (Colobus guereza kikuyuensis, Papio hamadryas, Homo sapiens, Sus scrofa domesticus, Capra hircus, Canis lupus familiaris, Bos taurus, Mus domesticus, and Myodes glareolus) from Spain. The data show that Trichuris spp. could be divided in three clades: Clade 1 = T. arvicolae, T. muris, and T.vulpis; Clade 2 = T. suis, T. colobae, T. trichiura, and T. spp. ex Papio hamadryas; Clade 3 = T. discolor, T. ovis, and T. skrjabini [69].
2.3. Immunomodulatory Effect of Trichuris spp.
Trichuris spp. has been used in the treatment of gastrointestinal autoimmune diseases, such as inflammatory bowel disease, Crohn’s disease, and ulcerative colitis [70,71,72]. Trichuris suis (pig whipworm) had been experimentally given to humans with no overt sign of gastrointestinal illness. The eggs produced from the feces remained constant, and only a low percentage of these eggs embryonated in vitro [21]. Some authors also noted that treatment of patients with inflammatory bowel disease, ulcerative colitis, and Crohn’s disease with Trichuris suis showed improvement in gastrointestinal signs, and in the management of disease the subjects were given ova every three weeks [70,71,72]. Surprisingly, Kradin et al. [22] noted that a patient that underwent treatment for Crohn’s disease using T. suis had adult worms beneath the ileocecal mucosal epithelium. This case does raise concerns about persistent infection from T. suis in man [22].
Further work was done on the use of excretory secretory products of T. suis in rats [73]. The investigation of the use of excretory products of T. suis in swine epithelium cells was used as a model to be used in humans. It was noted that the excretory secretory products (ESPs) elicited the production of interleukin (IL)-6 and IL-10, which have been identified as anti-inflammatory cytokines that inhibit Th-1 responses. This proved that ESPs from T. suis have immunomodulatory effects and can be used as candidates in the treatment of inflammatory bowel disease [73]. The use of ESPs from T. suis may be safer than the actual treatment with ova.
Subsequent research was done on the immunomodulatory and immunogenic effects of the proteins and ESPs of Trichuris trichiura and Trichuris muris [74,75,76]. Proteins were analyzed from adult worm extract and fragments of T. trichiura. These extracts and fragments were placed in cell cultures of human peripheral blood monocytes, and elicited the production of IL-10, IL-12, and TNF-α. Some fractions showed the inhibition of IL-5 production. The downregulation of IL-5 is a feature of a Th-2 response [74]. Santos et al. [74] concluded that protein fractions of T. trichiura can be used in the treatment and prevention of allergic and autoimmune diseases. Immunogenic research was also conducted on the ESPs of T. muris, and specific immunogenic proteins were identified. The structure of one such protein was Tm16, which was characterized and could be used in the production of a vaccine [75]. Shears et al. [76] noted that ESPs from T. muris elicited production of IL-9 and IL-13 when inoculated into rats. Eleven immunogenic proteins from the ESP of T. muris were also identified, and these could be used in the production of a vaccine [76]. Recent studies show that there is tremendous potential for Trichuris in human autoimmune disease, as well as vaccine development in rural countries where trichuriasis infections are prevalent.
This entry is adapted from the peer-reviewed paper 10.3390/vetsci8020015
This entry is offline, you can click here to edit this entry!