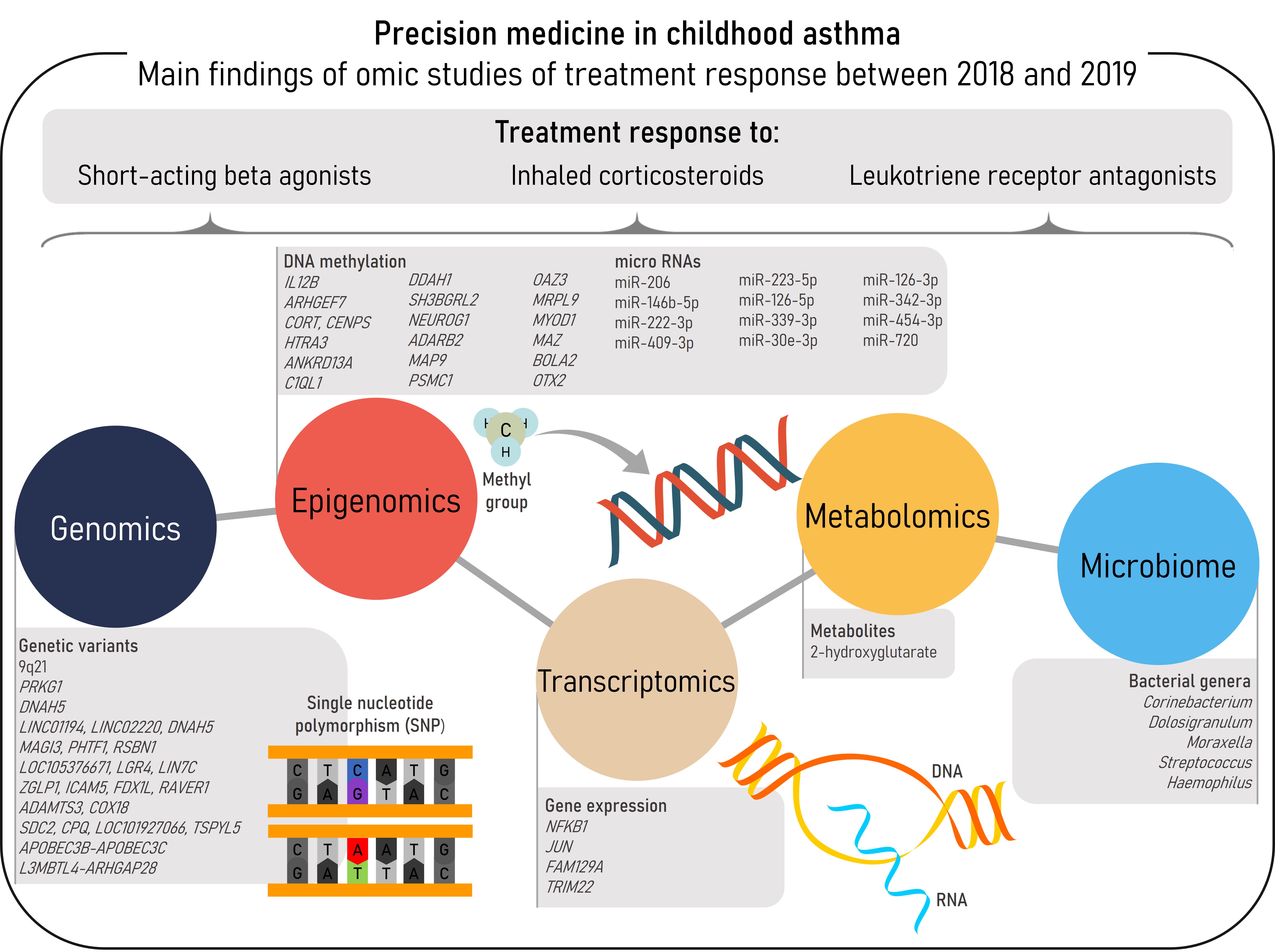
Asthma is a complex and multifactorial respiratory disease with a high prevalence in the pediatric population. Variation in treatment response to asthma therapies has been described among patients, and difficult-to-treat asthma carries both high healthcare and socioeconomic burden to the patients and society. Omic studies can be used to discover the molecular mechanisms underlying asthma susceptibility and treatment response, contributing to a better knowledge and definition of asthma pathogenesis and therefore, to the development of precision medicine. This entry aims to summarize the recent findings of omic studies of treatment response in childhood asthma. Between 2018-2019 a total of 13 omic studies has been performed involving genomics, epigenomics, transcriptomics, metabolomics, and the microbiome. These have been focused on the response to three common asthma medications: short-acting beta agonists, inhaled corticosteroids, and leukotriene receptor antagonists. Novel associations of different biomarkers with asthma treatment response have been described. However, stronger evidence and more consistent results are required to implement these molecular biomarkers into clinical practice by establishing the most appropriate therapy for each patient.
- asthma
- genomics
- transcriptomics
- epigenomics
- metabolomics
- microbiome
- omics
- inhaled corticosteroid
- short-acting beta agonist
- leukotriene receptor antagonist
Introduction
Pharmacogenomics
| rsID/ Chr. Band |
Chr:Positiona | Gene/ Nearest Gene |
Effect Allele | Effect | p-value | Reference |
|---|---|---|---|---|---|---|
| SABA response | ||||||
| rs73650726 | 9:85152666 | 9q21 | A | β=−3.8 | 7.69×10−9 | [12] |
| rs7903366 | 10:53689774 | PRKG1 | T | β=1.23 | 3.94×10−8 | |
| rs7070958 | 10:53691116 | PRKG1 | A | β=−1.24 | 4.09×10−8 | |
| rs7081864 | 10:53690331 | PRKG1 | A | β=1.23 | 4.94×10−8 | |
| rs17834628 | 5:12978566 | LINC01194 LINC02220 DNAH5 | A | OR=1.67 | 1.18×10−8 | [11] |
| rs35661809 | 5:12968341 | LINC01194 LINC02220 DNAH5 | G | OR=1.59 | 3.33×10−8 | |
| 1p13.2 | 1:114177000-1:114178000 | MAGI3 PHTF1 RSBN1 | NA | NA | 4.40×10−9 | |
| 11p14.1 | 11:27507000-11:27508000 | LOC105376671 LGR4 LIN7C | NA | NA | 6.59×10−9 | |
| 19p13.2 | 19:10424000-19:10425000 | ZGLP1 ICAM5 FDX1L RAVER1 | NA | NA | 3.12×10−11 | |
| 4q13.3 | 4:73478000-4:73479000 | ADAMTS3 COX18 | NA | NA | 6.25×10−8 | |
| 8q22.1 | 8:97926000-8:97927000 | SDC2 CPQ LOC101927066 TSPYL5 | NA | NA | 1.32×10−7 | |
| ICS response | ||||||
| rs5995653 | 22:39404249 | APOBEC3B-APOBEC3C | A | OR=0.70 | 3.31×10−7 | [13] |
| rs62081416 | 18:6605442 | L3MBTL4-ARHGAP28 | A | OR=2.44 | 1.57×10−5 | |
| aPosition based on GRCh37/hg19 build. rsID: reference SNP cluster ID; Chr: chromosome; SABA: short-acting beta agonist; ICS: inhaled corticosteroid; OR: odds ratio; NA: not available. | ||||||
Epigenomics
| CpG | Chromosome:Positiona | Gene/Nearest Gene | β | p-value | Reference |
|---|---|---|---|---|---|
| ICS response | |||||
| cg00066816 | 5:158758353 | IL12B | −3.101 | 0.002 | [24] |
| cg00557354 | 13:111767899 | ARHGEF7 | −3.490 | 0.001 | |
| cg04256470 | 1:10510465 | CORT CENPS | 3.620 | <0.001 | |
| cg09495977 | 4:8271507 | HTRA3 | −2.420 | 0.017 | |
| cg12333095 | 12:110437035 | ANKRD13A | −3.485 | 0.001 | |
| cg13818573 | 17:43045372 | C1QL1 | −3.596 | <0.001 | |
| cg21589280 | 1:85930152 | DDAH1 | −3.063 | 0.003 | |
| cg03080985 | 6:80340683 | SH3BGRL2 | −3.077 | 0.003 | |
| cg04330449 | 5:134871166 | NEUROG1 | −2.646 | 0.009 | |
| cg05307923 | 10:1779667 | ADARB2 | −2.577 | 0.011 | |
| cg08724517 | 4:156298205 | MAP9 | 2.951 | 0.004 | |
| cg11665562 | 14:90723462 | PSMC1 | −3.250 | 0.001 | |
| cg14269514 | 1:151736130 | OAZ3 MRPL9 | −3.112 | 0.002 | |
| cg24322623 | 11:17740431 | MYOD1 | −2.964 | 0.004 | |
| cg27254601 | 16:29817104 | MAZ BOLA2 | 3.598 | 0.0005 | [25] |
| cg15607672 | 14:57277228 | OTX2 | 2.123 | 0.0363 | |
| aPosition based on GRCh37/hg19 build. DNAm: DNA methylation; ICS: inhaled corticosteroid. | |||||
| miRNA | Odds Ratio | p-value |
|---|---|---|
| miR-206 | 0.60 | 0.004 |
| miR-146b-5p | 0.66 | 0.007 |
| miR-222-3p | 0.70 | 0.02 |
| miR-409-3p | 0.73 | 0.02 |
| miR-223-5p | 0.62 | 0.02 |
| miR-126-5p | 0.68 | 0.03 |
| miR-339-3p | 0.72 | 0.03 |
| miR-30e-3p | 0.70 | 0.03 |
| miR-126-3p | 0.74 | 0.03 |
| miR-342-3p | 0.80 | 0.04 |
| miR-454-3p | 0.77 | 0.04 |
| miR-720 | 0.71 | 0.046 |
| miRNA: micro ribonucleic acid. | ||
Transcriptomics
Metabolomics
Microbiome
Conclusions
References
- The Global Asthma Report 2018; Global Asthma Network: Auckland, New Zealand, 2018; ISBN 978-0-473-29125-9\r978-0-473-29126-6.
- Global Initiative for Asthma. Global Strategy for Asthma Management and Prevention; Global Initiative for Asthma: Fontana, WI, USA, 2019. [Google Scholar]
- Guilbert, T.W.; Bacharier, L.B.; Fitzpatrick, A.M. Severe Asthma in Children. J. Allergy Clin. Immunol. Pract. 2014, 2, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.; Pereira, A.M.; Morais-Almeida, M. Asthma Costs and Social Impact. Asthma Res. Pract. 2017, 3, 1. [Google Scholar] [CrossRef]
- Tyler, S.R.; Bunyavanich, S. Leveraging -Omics for Asthma Endotyping. J. Allergy Clin. Immunol. 2019, 144, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Visscher, P.M.; Wray, N.R.; Zhang, Q.; Sklar, P.; McCarthy, M.I.; Brown, M.A.; Yang, J. 10 Years of GWAS Discovery: Biology, Function, and Translation. Am. J. Hum. Genet. 2017, 101, 5–22. [Google Scholar] [CrossRef]
- Petersen, B.S.; Fredrich, B.; Hoeppner, M.P.; Ellinghaus, D.; Franke, A. Opportunities and Challenges of Whole-Genome and -Exome Sequencing. BMC Genet. 2017, 18, 14. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Luis, E.; Hernandez-Pacheco, N.; Vijverberg, S.J.; Flores, C.; Pino-Yanes, M. Role of Genomics in Asthma Exacerbations. Curr. Opin. Pulm. Med. 2019, 25, 101–112. [Google Scholar] [CrossRef]
- García-Menaya, J.M.; Cordobés-Durán, C.; García-Martín, E.; Agúndez, J.A.G. Pharmacogenetic Factors Affecting Asthma Treatment Response. Potential Implications for Drug Therapy. Front. Pharmacol. 2019, 10, 520. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Pacheco, N.; Flores, C.; Oh, S.S.; Burchard, E.G.; Pino-Yanes, M. What Ancestry Can Tell Us About the Genetic Origins of Inter-Ethnic Differences in Asthma Expression. Curr. Allergy Asthma Rep. 2016, 16, 53. [Google Scholar] [CrossRef] [PubMed]
- Mak, A.C.Y.; White, M.J.; Eckalbar, W.L.; Szpiech, Z.A.; Oh, S.S.; Pino-Yanes, M.; Hu, D.; Goddard, P.; Huntsman, S.; Galanter, J.; et al. Whole-Genome Sequencing of Pharmacogenetic Drug Response in Racially Diverse Children with Asthma. Am. J. Respir. Crit. Care Med. 2018, 197, 1552–1564. [Google Scholar] [CrossRef] [PubMed]
- Spear, M.L.; Hu, D.; Pino-Yanes, M.; Huntsman, S.; Eng, C.; Levin, A.M.; Ortega, V.E.; White, M.J.; McGarry, M.E.; Thakur, N.; et al. A Genome-Wide Association and Admixture Mapping Study of Bronchodilator Drug Response in African Americans with Asthma. Pharmacogenomics J. 2019, 19, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Pacheco, N.; Farzan, N.; Francis, B.; Karimi, L.; Repnik, K.; Vijverberg, S.J.; Soares, P.; Schieck, M.; Gorenjak, M.; Forno, E.; et al. Genome-Wide Association Study of Inhaled Corticosteroid Response in Admixed Children with Asthma. Clin. Exp. Allergy 2019, 49, 789–798. [Google Scholar] [CrossRef]
- Kanazawa, H.; Hirata, K.; Yoshikawa, J. Nitrogen Oxides Reduce Albuterol-Induced Bronchodilation in Patients with Bronchial Asthma. Respiration 2002, 69, 490–495. [Google Scholar] [CrossRef]
- Ricciardolo, F.L.M. Multiple Roles of Nitric Oxide in the Airways. Thorax 2003, 58, 175–182. [Google Scholar] [CrossRef]
- Ferreira, M.A.R.; Matheson, M.C.; Duffy, D.L.; Marks, G.B.; Hui, J.; Le Souëf, P.; Danoy, P.; Baltic, S.; Nyholt, D.R.; Jenkins, M.; et al. Identification of IL6R and Chromosome 11q13.5 as Risk Loci for Asthma. Lancet 2011, 378, 1006–1014. [Google Scholar] [CrossRef]
- Kichaev, G.; Bhatia, G.; Loh, P.R.; Gazal, S.; Burch, K.; Freund, M.K.; Schoech, A.; Pasaniuc, B.; Price, A.L. Leveraging Polygenic Functional Enrichment to Improve GWAS Power. Am. J. Hum. Genet. 2019, 104, 65–75. [Google Scholar] [CrossRef]
- Lutz, S.M.; Cho, M.H.; Young, K.; Hersh, C.P.; Castaldi, P.J.; McDonald, M.L.; Regan, E.; Mattheisen, M.; DeMeo, D.L.; Parker, M.; et al. A Genome-Wide Association Study Identifies Risk Loci for Spirometric Measures among Smokers of European and African Ancestry. BMC Genet. 2015, 16, 138. [Google Scholar] [CrossRef]
- Lee, J.H.; McDonald, M.L.; Cho, M.H.; Wan, E.S.; Castaldi, P.J.; Hunninghake, G.M.; Marchetti, N.; Lynch, D.A.; Crapo, J.D.; Lomas, D.A.; et al. DNAH5 Is Associated with Total Lung Capacity in Chronic Obstructive Pulmonary Disease. Respir. Res. 2014, 15, 97. [Google Scholar] [CrossRef]
- Ramasamy, A.; Curjuric, I.; Coin, L.J.; Kumar, A.; McArdle, W.L.; Imboden, M.; Leynaert, B.; Kogevinas, M.; Schmid-Grendelmeier, P.; Pekkanen, J.; et al. A Genome-Wide Meta-Analysis of Genetic Variants Associated with Allergic Rhinitis and Grass Sensitization and Their Interaction with Birth Order. J. Allergy Clin. Immunol. 2011, 128, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Forno, E.; Celedón, J.C. Epigenomics and Transcriptomics in the Prediction and Diagnosis of Childhood Asthma: Are We There Yet? Front. Pediatr. 2019, 7, 115. [Google Scholar] [CrossRef] [PubMed]
- Fazzari, M.J.; Greally, J.M. Introduction to Epigenomics and Epigenome-Wide Analysis. Methods Mol. Biol. 2010, 620, 243–265. [Google Scholar] [CrossRef]
- Kronfol, M.M.; Dozmorov, M.G.; Huang, R.; Slattum, P.W.; McClay, J.L. The Role of Epigenomics in Personalized Medicine. Expert Rev. Precis. Med. Drug Dev. 2017, 2, 33–45. [Google Scholar] [CrossRef]
- Wang, A.L.; Gruzieva, O.; Qiu, W.; Kebede Merid, S.; Celedón, J.C.; Raby, B.A.; Söderhäll, C.; DeMeo, D.L.; Weiss, S.T.; Melén, E.; et al. DNA Methylation Is Associated with Inhaled Corticosteroid Response in Persistent Childhood Asthmatics. Clin. Exp. Allergy 2019, 49, 1225–1234. [Google Scholar] [CrossRef]
- Wang, A.L.; Qiu, W.; Demeo, D.L.; Raby, B.A.; Weiss, S.T.; Tantisira, K.G. DNA Methylation Is Associated with Improvement in Lung Function on Inhaled Corticosteroids in Pediatric Asthmatics. Pharmacogenet. Genom. 2019, 29, 65–68. [Google Scholar] [CrossRef]
- Szabo, S.J.; Jacobson, N.G.; Dighe, A.S.; Gubler, U.; Murphy, K.M. Developmental Commitment to the Th2 Lineage by Extinction of IL-12 Signaling. Immunity 1995, 2, 665–675. [Google Scholar] [CrossRef]
- Naseer, T.; Minshall, E.M.; Leung, D.Y.M.; Laberge, S.; Ernst, P.; Martin, R.J.; Hamid, Q. Expression of IL-12 and IL-13 MRNA in Asthma and Their Modulation in Response to Steroid Therapy. Am. J. Respir. Crit. Care Med. 1997, 155, 845–851. [Google Scholar] [CrossRef]
- Keane-Myers, A.; Wysocka, M.; Trinchieri, G.; Wills-Karp, M. Resistance to Antigen-Induced Airway Hyperresponsiveness Requires Endogenous Production of IL-12. J. Immunol. 1998, 161, 919–926. [Google Scholar]
- Giordano, R.; Picu, A.; Bonelli, L.; Broglio, F.; Prodam, F.; Grottoli, S.; Muccioli, G.; Ghigo, E.; Arvat, E. The Activation of Somatostatinergic Receptors by Either Somatostatin-14 or Cortistatin-17 Often Inhibits ACTH Hypersecretion in Patients with Cushing’s Disease. Eur. J. Endocrinol. 2007, 157, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rey, E.; Pedreño, M.; Delgado-Maroto, V.; Souza-Moreira, L.; Delgado, M. Lulling Immunity, Pain, and Stress to Sleep with Cortistatin. Ann. N. Y. Acad. Sci. 2015, 1351, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Frey, A.G.; Palenchar, D.J.; Wildemann, J.D.; Philpott, C.C. A Glutaredoxin-BolA Complex Serves as an Iron-Sulfur Cluster Chaperone for the Cytosolic Cluster Assembly Machinery. J. Biol. Chem. 2016, 291, 22344–22356. [Google Scholar] [CrossRef] [PubMed]
- Bigler, J.; Boedigheimer, M.; Schofield, J.P.R.; Skipp, P.J.; Corfield, J.; Rowe, A.; Sousa, A.R.; Timour, M.; Twehues, L.; Hu, X.; et al. A Severe Asthma Disease Signature from Gene Expression Profiling of Peripheral Blood from U-BIOPRED Cohorts. Am. J. Respir. Crit. Care Med. 2017, 195, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Astle, W.J.; Elding, H.; Jiang, T.; Allen, D.; Ruklisa, D.; Mann, A.L.; Mead, D.; Bouman, H.; Riveros-Mckay, F.; Kostadima, M.A.; et al. The Allelic Landscape of Human Blood Cell Trait Variation and Links to Common Complex Disease. Cell 2016, 167, 1415–1429.e19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Biagini Myers, J.M.; Yadagiri, V.K.; Ulm, A.; Chen, X.; Weirauch, M.T.; Khurana Hershey, G.K.; Ji, H. Nasal DNA Methylation Differentiates Corticosteroid Treatment Response in Pediatric Asthma: A Pilot Study. PLoS ONE 2017, 12, e0186150. [Google Scholar] [CrossRef] [PubMed]
- Kho, A.T.; McGeachie, M.J.; Moore, K.G.; Sylvia, J.M.; Weiss, S.T.; Tantisira, K.G. Circulating MicroRNAs and Prediction of Asthma Exacerbation in Childhood Asthma. Respir. Res. 2018, 19, 128. [Google Scholar] [CrossRef]
- Kho, A.T.; Sharma, S.; Davis, J.S.; Spina, J.; Howard, D.; McEnroy, K.; Moore, K.; Sylvia, J.; Qiu, W.; Weiss, S.T.; et al. Circulating MicroRNAs: Association with Lung Function in Asthma. PLoS ONE 2016, 11, e0157998. [Google Scholar] [CrossRef]
- Yeh, Y.L.; Su, M.W.; Chiang, B.L.; Yang, Y.H.; Tsai, C.H.; Lee, Y.L. Genetic Profiles of Transcriptomic Clusters of Childhood Asthma Determine Specific Severe Subtype. Clin. Exp. Allergy 2018, 48, 1164–1172. [Google Scholar] [CrossRef]
- Qiu, W.; Guo, F.; Glass, K.; Yuan, G.C.; Quackenbush, J.; Zhou, X.; Tantisira, K.G. Differential Connectivity of Gene Regulatory Networks Distinguishes Corticosteroid Response in Asthma. J. Allergy Clin. Immunol. 2018, 141, 1250–1258. [Google Scholar] [CrossRef] [PubMed]
- Beste, M.T.; Pfäffle-Doyle, N.; Prentice, E.A.; Morris, S.N.; Lauffenburger, D.A.; Isaacson, K.B.; Griffith, L.G. Endometriosis: Molecular Network Analysis of Endometriosis Reveals a Role for c-Jun-Regulated Macrophage Activation. Sci. Transl. Med. 2014, 6, 222ra16. [Google Scholar] [CrossRef] [PubMed]
- McGeachie, M.J.; Clemmer, G.L.; Hayete, B.; Xing, H.; Runge, K.; Wu, A.C.; Jiang, X.; Lu, Q.; Church, B.; Khalil, I.; et al. Systems Biology and in Vitro Validation Identifies Family with Sequence Similarity 129 Member A (FAM129A) as an Asthma Steroid Response Modulator. J. Allergy Clin. Immunol. 2018, 142, 1479–1488.e12. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Wang, J.; Liu, J.; Huang, Y.; Zhou, Y.; Yang, S.; Zhang, W.; Yang, M.; Zhang, H. Niban Protein Regulates Apoptosis in HK-2 Cells via Caspase-Dependent Pathway. Ren. Fail. 2019, 41, 455–466. [Google Scholar] [CrossRef]
- Katayama, S.; Hammar, K.S.; Krjutškov, K.; Einarsdottir, E.; Hedlin, G.; Kere, J.; Söderhäll, C. Acute Wheeze-Specific Gene Module Shows Correlation with Vitamin D and Asthma Medication. Eur. Respir. J. 2019, 55, 1901330. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Lian, Q.; Sun, B. Interferons Command Trim22 to Fight against Viruses. Cell. Mol. Immunol. 2017, 14, 794–796. [Google Scholar] [CrossRef]
- Pité, H.; Morais-Almeida, M.; Rocha, S.M. Metabolomics in Asthma: Where Do We Stand? Curr. Opin. Pulm. Med. 2018, 24, 94–103. [Google Scholar] [CrossRef]
- Neerincx, A.H.; Vijverberg, S.J.H.; Bos, L.D.J.; Brinkman, P.; van der Schee, M.P.; de Vries, R.; Sterk, P.J.; Maitland-van der Zee, A.-H. Breathomics from Exhaled Volatile Organic Compounds in Pediatric Asthma. Pediatr. Pulmonol. 2017, 52, 1616–1627. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, S.H.; Kim, J.H.; Hwang, S.; Yoo, H.J. Understanding Metabolomics in Biomedical Research. Endocrinol. Metab. 2016, 31, 7–16. [Google Scholar] [CrossRef]
- Kelly, R.S.; Sordillo, J.E.; Lutz, S.M.; Avila, L.; Soto-Quiros, M.; Celedón, J.C.; McGeachie, M.J.; Dahlin, A.; Tantisira, K.; Huang, M.; et al. Pharmacometabolomics of Bronchodilator Response in Asthma and the Role of Age-Metabolite Interactions. Metabolites 2019, 9, 179. [Google Scholar] [CrossRef]
- Kelly, R.S.; Chawes, B.L.; Blighe, K.; Virkud, Y.V.; Croteau-Chonka, D.C.; McGeachie, M.J.; Clish, C.B.; Bullock, K.; Celedón, J.C.; Weiss, S.T.; et al. An Integrative Transcriptomic and Metabolomic Study of Lung Function in Children With Asthma. Chest 2018, 154, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Paulenda, T.; Draber, P. The Role of ORMDL Proteins, Guardians of Cellular Sphingolipids, in Asthma. Allergy Eur. J. Allergy Clin. Immunol. 2016, 71, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Toncheva, A.A.; Potaczek, D.P.; Schedel, M.; Gersting, S.W.; Michel, S.; Krajnov, N.; Gaertner, V.D.; Klingbeil, J.M.; Illig, T.; Franke, A.; et al. Childhood Asthma Is Associated with Mutations and Gene Expression Differences of ORMDL Genes That Can Interact. Allergy Eur. J. Allergy Clin. Immunol. 2015, 70, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Ono, J.G.; Worgall, T.S.; Worgall, S. 17q21 Locus and ORMDL3: An Increased Risk for Childhood Asthma. Pediatr. Res. 2014, 75, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Weinstock, G.M. Genomic Approaches to Studying the Human Microbiota. Nature 2012, 489, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, R.; Rani, A.; Metwally, A.; McGee, H.S.; Perkins, D.L. Analysis of the Microbiome: Advantages of Whole Genome Shotgun versus 16S Amplicon Sequencing. Biochem. Biophys. Res. Commun. 2016, 469, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Jackson, D.; Bacharier, L.B.; Mauger, D.; Boushey, H.; Castro, M.; Durack, J.; Huang, Y.; Lemanske, R.F.; Storch, G.A.; et al. The Upper-Airway Microbiota and Loss of Asthma Control among Asthmatic Children. Nat. Commun. 2019, 10, 5714. [Google Scholar] [CrossRef]
- Durack, J.; Huang, Y.J.; Nariya, S.; Christian, L.S.; Mark Ansel, K.; Beigelman, A.; Castro, M.; Dyer, A.M.; Israel, E.; Kraft, M.; et al. Bacterial Biogeography of Adult Airways in Atopic Asthma. Microbiome 2018, 6, 104. [Google Scholar] [CrossRef]
- Chung, K.F. Airway Microbial Dysbiosis in Asthmatic Patients: A Target for Prevention and Treatment? J. Allergy Clin. Immunol. 2017, 139, 1071–1081. [Google Scholar] [CrossRef]
- Depner, M.; Ege, M.J.; Cox, M.J.; Dwyer, S.; Walker, A.W.; Birzele, L.T.; Genuneit, J.; Horak, E.; Braun-Fahrländer, C.; Danielewicz, H.; et al. Bacterial Microbiota of the Upper Respiratory Tract and Childhood Asthma. J. Allergy Clin. Immunol. 2017, 139, 826–834.e13. [Google Scholar] [CrossRef]
- McCauley, K.; Durack, J.; Valladares, R.; Fadrosh, D.W.; Lin, D.L.; Calatroni, A.; LeBeau, P.K.; Tran, H.T.; Fujimura, K.E.; LaMere, B.; et al. Distinct Nasal Airway Bacterial Microbiotas Differentially Relate to Exacerbation in Pediatric Patients with Asthma. J. Allergy Clin. Immunol. 2019, 144, 1187–1197. [Google Scholar] [CrossRef]
- Zhou, Y.; Gao, H.; Mihindukulasuriya, K.A.; Rosa, P.S.L.; Wylie, K.M.; Vishnivetskaya, T.; Podar, M.; Warner, B.; Tarr, P.I.; Nelson, D.E.; et al. Biogeography of the Ecosystems of the Healthy Human Body. Genome Biol. 2013, 14, R1. [Google Scholar] [CrossRef]
This entry is adapted from the peer-reviewed paper 10.3390/ijms21082908