1. Avian Influenza A Virus Infection Transmission to Humans
AIVs are globally challenging due to widespread circulation. They are differentiated into low- and high-pathogenic viral strains. Low-pathogenic viral strains only cause mild illness in domestic birds. Conversely, highly pathogenic avian phenotypes, such as H5N1, H5N8, and H7N9, may cause organ damage, affect the respiratory tract, and determine up to 100% bird mortality [
10,
11]. Generally, highly pathogenic AIVs emerge from low-pathogenic viruses, acquiring cleavage site insertions that promote systemic infections [
12].
AIVs show the potential to be episodically transmitted from birds to other animals and humans by direct contact or through an intermediate host. AIV infections in humans range in severity from no symptoms or mild illness (fever, conjunctivitis, and mild influenza-like upper respiratory symptoms) to severe diseases (pneumonia, acute respiratory distress syndrome, multi-organ dysfunction, and encephalopathy) and death; less common infection signs include diarrhea, nausea, vomiting, and seizures [
6]. Most serious illnesses and high mortality rates have been associated with the Asian lineages H7N9 and H5N1 [
3,
13].
Although AIVs spread from one infected person to close contacts, prolonged and unprotected contact is rare and generally limited to a few people, there is a global concern that highly pathogenic AIVs (H5/H7/H9/H10) are adapting to other species by acquiring mutations in animals or humans that support their cross-species transmission [
5,
14,
15,
16]. Notably, the segmented RNA genome allows IAVs from different species to mix their genes by reassortment, generating new viruses. Important, adaptation processes of AIVs are needed for their efficient transmission and replication in mammal hosts. This aspect first includes the receptor–binding specificity switch of HA to the mammalian sialic acid 2,6 galactose-type receptor [
17]. Moreover, the presence of specific mutations in the viral polymerase complex is considered an important host range determinant [
18,
19]. In addition, the NA of AIVs contains a specific sialic acid-binding site that is absent in human IAV, affecting the catalytic enzyme activity [
20].
Since 1996, 11 subtypes of AIVs have been assessed to spread directly from infected birds to humans. Importantly, infected humans can transmit influenza viruses to other animals (pigs, poultry, cats, and dogs). This phenomenon is called reverse zoonosis and may contribute to the further diffusion of influenza viruses [
21].
The highly pathogenic H5N1 AIV subtype emerged in China in 1996, establishing sustained transmission in domestic poultry [
22]. From 1997, H5N1 human infections occurred in Asian countries with high mortality levels [
23,
24]; the results of serological assays demonstrated the human-to-human virus spread [
25,
26]. In a second distribution wave in 2003, the H5N1 virus was disseminated in Asia and Europe through migratory birds [
27]. From 2003, several human H5N1 infection episodes were reported worldwide, and, up to 2023, the WHO documented 457 fatalities [
28]. Since 2021, the H5N1 virus caused several infections in wild carnivores, mink farms, and marine mammals [
29]. The wider range of H5N1 infections and the involvement of different animal species have favored the emergence of new and potentially more virulent variants. Importantly, continuous genetic variability has been observed for the H5N1 virus across various geographic regions as well as increased virulence as it evolves [
22,
30,
31,
32,
33,
34]. In 2014, a fatal human infection by the novel H5N6 AIV was also reported [
35].
H9N2 AIV has been circulating in poultry since 1994 and is occasionally transmitted to humans [
36,
37,
38]. Importantly, the expanded receptor specificity of H9N2 has raised concerns as a possible source of a novel human influenza virus [
39,
40].
From 2003 to 2017, sporadic human infections caused by H5N1, H7N7, and H7N9 AIVs have been observed in different European and Asian countries [
41,
42,
43,
44,
45,
46]. Since 2013, newly emerging AIVs have frequently crossed the species barrier to infect humans, causing several fatal infections [
47,
48,
49,
50,
51].
Overall, it has to be considered that both continuous influenza virus genetic evolution and the actual environment of human activities may facilitate the occurrence of zoonoses with a high transmissibility level, as evidenced by the recent SARS-CoV-2 pandemic. Moreover, highly pathogenic AIVs have never circulated extensively among humans; therefore, the lack of pre-existing immunity poses the human population with a high risk of severe influenza occurrence.
2. The Cellular Cytoskeleton
The cytoskeleton of eukaryotic cells is a well-organized and highly dynamic filamentous network that radiates through the cell and primarily comprises three filament types that function co-ordinately: actin filaments (microfilaments), microtubules, and intermediate filaments [
52]. Septins are a family of GTPases and are considered the fourth cytoskeleton component [
53]. Cellular motor proteins, such as dynein, kinesin, myosin, and other accessory proteins, represent a structurally and functionally diverse family of molecules that actively concur with cytoskeletal functions, facilitating the capture and transport of a variety of cargo along cytoskeletal networks.
A complex cytoskeletal network connects the plasma membrane to the nucleus and carries out diverse roles [
54]. Specifically, the cytoskeleton contributes to cell morphology regulation, cell migration, apoptosis, cell differentiation, and cell division [
52,
55,
56,
57,
58,
59]. Importantly, all cytoskeletal components can swiftly adapt to both external and intracellular stimuli, undergoing rapid and continuous structural modifications [
60].
Microfilaments are thin and fibrous structures responsible for cytoplasmic streaming and represent the major structural component of the cell. They concentrate under the plasma membrane, are involved in cell division, maintain the plasma membrane structure, and participate in intracellular trafficking and cell motility. Several actin-binding proteins connect actin filaments, while other actin-binding proteins allow the interaction of actin with specific cytoskeleton components [
61]. Microfilaments are considered the most dynamic cytoskeletal networks, as they can undergo rapid and significant depolymerization when cell deformation and movement are required [
60,
62]. Specifically, cortical actin is primarily implied in events related to the presentation of cell membrane molecules, endocytosis, and viral entry/exit mechanisms [
63], while nuclear actin regulates chromatin remodeling and gene transcription mechanisms [
64].
Microtubules are highly dynamic polymers of tubulin subjected to rapid cycles of polymerization and subsequent depolymerization, depending on the requirements of the cell [
65]. Microtubules radiate from the centrosome, the microtubule-organizing center of eukaryotic cells [
66]. They are involved in cell motility, signal transduction, the subcellular distribution of organelles, and intracellular transport [
67]. Several microtubule-binding proteins regulate microtubule dynamics [
68]. Both actin and microtubules form polarized filaments with growing plus-ends pointed toward the plasma membrane, allowing the ATP-mediated directional transport of cargo such as vesicles and organelles. Specifically, actin is generally involved in the short-range transport near the cytoplasmic membrane, while microtubules are for the long-range intracellular transport.
The largest gene family encodes the intermediate filaments (IFs). IFs are found in the cytoplasm and are adjacent to the inner face of the nuclear envelope. Comprised of more than 70 components, they show a remarkable diversity, and provide mechanical strength to animal cells, maintaining the cell shape and tension, and protecting the genome. Major IFs are classified into five types, based on their structural composition and origin: keratin, desmin, glial fibrillary acidic protein, vimentin, and neurofilament protein [
69]. Keratins are the main components of the cytoskeleton in epithelial and mesenchymal cells. Keratins are classified into two groups, based on differential immunogenicity properties. They provide structural support to cells and counteract the effects of physical stress on cell integrity. Vimentin is assembled into major cytoskeletal systems in cells of mesenchymal and ectodermal origin. Recent evidence has shown that vimentin and filamentous actin form interpenetrating networks in the cell cortex and vimentin participates in the regulation of actin dynamics [
70]. The intracellular distribution of vimentin allows the structural maintenance of cell organelles. Moreover, vimentin is an integrator of cellular mechanical processes, promptly responding to cellular stress [
71].
Lamins are nuclear IFs and the principal component of the nuclear lamina, a mesh of proteins inside the inner nuclear membrane [
72]. Lamins maintain the structural integrity of the nucleus and anchor the chromatin and nuclear pore complexes to the nuclear periphery [
73]. In association with actin and actin motor proteins, these proteins mainly constitute the nuclear cytoskeleton [
74]. Of note, they are involved in key roles, such as the regulation of nuclear morphology, stability, gene expression, and transmitting and deciphering mechanical stimuli into physiological responses.
Overall, the cytoskeletal scaffold regulates numerous aspects of cell biology, undergoing rapid and continuous structural changes to satisfy the requirements of the cell in both physiological and pathological conditions. Viruses represent an excellent tool to study the regulatory mechanisms of the cellular cytoskeleton.
Viruses are obligate intracellular parasites that depend strictly on the host cell machinery to perform their successful replication [
52,
75,
76,
77]. Specifically, the cytoskeleton provides a scaffold for viral entry, sub-cellular trafficking through the dense cytosol, replication, assembly, and egression [
78]. Viruses can subvert and exploit physiological cytoskeletal functions to promote their replication, with varying effects, depending on the virus species involved. The complexity of the cellular cytoskeleton and the differences between the replication cycles of human viruses belonging to different families allow very different types of interactions, which remain poorly described. The contribution of the cytoskeleton in the replication cycle of viruses is critical to expand our knowledge of virus-coopted regulatory mechanisms, which could allow the discovery of potential therapeutic strategies.
To date, considerable research activities have attempted to understand the pathogenicity and mechanisms of transmission of AIVs, focusing on the influenza virus themselves. Therefore, a comprehensive understanding of the response of host factors during viral pathogenesis in mammals is needed. In this way, characterizing the complex molecular mechanisms linking the cell cytoskeleton to influenza virus infection provides insights into the main filament network alterations, which might be useful for successful virus replication and transmission.
3. The Involvement of the Cellular Cytoskeleton in Influenza Virus Infection
In influenza virus infection, several cytoskeletal proteins exert a key regulatory role during internalization, endosomal acidification, and virus intracellular transport mechanisms [
79,
80]. It has been shown that endosomes containing influenza viruses co-localize with microfilaments and undergo further movements, suggesting the intervention of myosin motor proteins [
81,
82]. Myosin is also involved when influenza virus entry occurs through micropinocytosis [
83]. In addition, keratin participates in the early cytoplasmic transport of influenza virus [
84].
In the second step, the influenza virus intracytoplasmic transport switches from microfilaments to microtubules and the associated dynein motor protein, allowing the release of viral ribonucleoproteins near the nucleus, the site of viral genome replication [
65,
85,
86]. It has been shown that intact microtubules promote both influenza virus entry to and exit from the host cell [
82,
87,
88].
Vimentin regulates the activity and intracellular transport of IAVs, also exerting critical roles in the host cell response [
80,
89]. Specifically, it has been assessed that vimentin might play an important role in the regulation of lipids during H9N2 replication, providing an important antiviral target against the influenza virus [
90]. Moreover, it has been shown that the integrity of IFs affects the release of H7N1 AIV progeny in mammalian cells, leading to decreased viral replication efficiency [
91].
In the late phases of infection, the microfilaments regulate the distribution of newly synthesized influenza virus proteins at the plasma membrane, with the parallel involvement of myosin, contributing to viral assembly and budding [
92,
93].
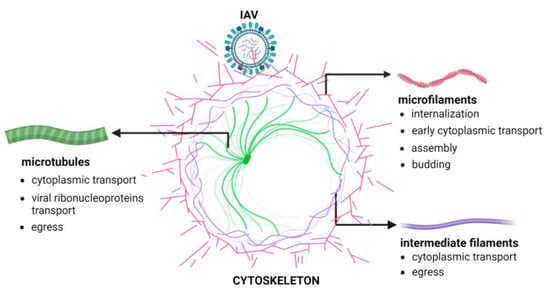
Figure 1 shows the phases of the IAV replication cycle requiring the active participation of the cellular cytoskeleton.
Figure 1. Cytoskeleton involvement during IAV replication. The Figure has been created with
BioRender.com (accessed on 6 March 2024).
The cell-dependent highly dynamic or polymerized state of the cytoskeleton and the associated regulatory proteins may regulate the exit of influenza virus infection. In this regard, mammalian Diaphanous-related formin-1, a Rho-effector protein generating linear actin filaments and regulating microtubule organization, represents a restriction factor that counteracts cytoskeleton dynamics during the early phases of IAV infection [
94]. In addition, it has been shown that highly polymerized/stable microfilaments and microtubules may restrict the early phases of IAV infection [
81,
91].
Notably, the influenza virus can modulate the dynamics of different cytoskeletal filaments by promoting their remodeling to support viral replication [
95]. In this regard, the IAV can subvert the structural organization of cytokeratin 8, enhancing its phosphorylation state and thereby promoting its replicative efficiency [
96]. Moreover, it has been assessed that the influenza virus stimulated the phosphorylation of cytoskeletal ezrin, radixin, and moesin, inducing structural cytoskeletal changes and permeability increases in pulmonary microvascular endothelial cells that favor its replication [
97].
These observations strongly support the contribution of specific components of the cellular cytoskeleton to the replication of influenza viruses. Their intervention has some degree of variability, depending on both the cell type and the viral strain involved, attesting to the uniqueness of the influenza virus–host cell relationship.
This entry is adapted from the peer-reviewed paper 10.3390/pathogens13030249