Plants experience stress facing unfavourable growth conditions. This is not only crucial in understanding the impact of unfavorable conditions on plant growth but also important for agriculture and food security. The unfavorable conditions may lead to yield loss of up to 70% for major crops, reflecting that average yields are limited to around 30% of their genetic potential [
1]. Abiotic stress caused by excesses or deficiencies in water, salt, light, temperature and nutrients, significantly lowers plant growth and productivity and even poses a threat to survival. Stressed plants have less-than-ideal environmental circumstances for development processes like cell division and expansion, which restricts plant advancement; dry-season stretch restrains plant growth since water is required for cell turgor, which applies weight on the cell development. Cold stress limits enzyme activities and other proteins, thus reducing plant growth.
Volatile organic compounds (VOCs), a type of phytochemical among secondary metabolites, are responsible for chemical signaling molecules that are involved in intra- and interplant communication, benefiting the health of both the emitter and adjacent beneficiary plants [
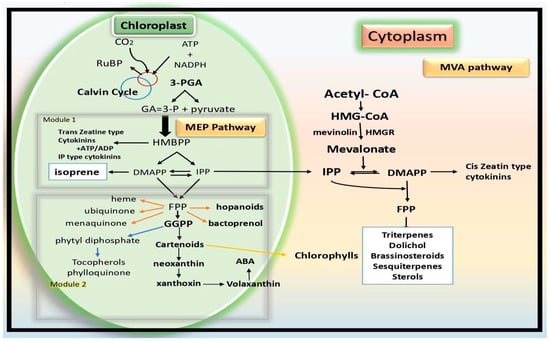
4]. Isoprene (C5H8), which is small and volatile, is produced at the chloroplast through the 2-C-methyl-
d-erythritol 4-phosphate (MEP) pathway. The synthesis of one isoprene molecule requires 14 NADPH and 21 ATP, indicating that it is energy- and carbon-intensive. The final step in the production of isoprene involves converting dimethylallyl diphosphate (DMADP) to isoprene, which is catalyzed by the enzyme known as “isoprene synthase” (IspS). In addition to producing hemiterpene isoprene, DMADP serves as a substrate for synthesizing several other important molecules such as monoterpenes, pigments including carotenoids, chlorophyll prenyl chains and cytokinins; hormones like abscisic acid (ABA); and pigments too [
5].
Plants use volatile organic compounds (VOCs) to communicate with other living things. Isoprene, monoterpenes and sesquiterpenes make up the largest class of volatile organic compounds (VOCs) emitted by plants terpenes. In plant–plant interactions, mono- and sesquiterpenes are well-known communication molecules. On the other hand, isoprene, the smallest and most often released terpene, is instead given a role in fighting abiotic stressors. Different isoprene-derived signaling molecules in plant systems are featured in the following sections.
2. Brassinosteroids
Brassinosteroids (BRs) are a class of signaling molecules with a steroidal skeleton with four rings. They play a noticeable role in the hormonal network that regulates numerous critical physiological processes in the plant life cycle. BRs play a role in male fertility, flowering and germination and control the lengthening of organs, the onset of senility and the plant’s capacity to withstand stresses like heat, salinity and water during the vegetative growth phase [
9]. BRs are steroid alcohols similar to other plant sterols. They share a structural resemblance with cholesterol, the main C27 animal sterol. BRs, along with other phytosterols, can have 27–29 carbon atoms, depending on how side chains are substituted (i.e., the type of C-24 alkyl substituent). According to [
10], C28 phytosterols, like BL, are the most abundant in plants. The entire steroid family requires the linear C30 hydrocarbon squalene, which insects cannot cyclize. In contrast, plants, animals and humans have access to the full sterol biosynthesis pathway. The cytoplasmic mevalonic acid pathway is the source of the isoprenoid building blocks IPP and DMAPP, which are used in BR synthesis.
The carbon skeleton of BR C27, an analog of BR C28 without the methyl group at C24 (norBRs), is identical to that of cholesterol, suggesting that they are synthesized from cholesterol via the same pathway as CS from CR. Additionally, the methylation of norCS at C24 in the presence of NADPH and S-adenosyl-l-methionine was shown to induce CS formation in cell-free enzymatic extracts of
Arabidopsis thaliana [
11].
Sitosterol, a naturally occurring C29 phytosterol, can form C29 BRs and 24-ethylBRs (also known as homoBRs) when combined with an enzyme extract from cell-free rice seedlings [
12]. The final biosynthetic product in this instance is homoCS, which is produced by homoTE and homoTY. Strangely, no homoBL was observed in plant tissues, even though similar processes described above result in the synthesis of either BL or its equivalent analog. Additionally, it appears that C28 demethylation of C29 BRs can produce C28 BRs, and this interconversion may enable plants to restore a BR bioactivity level that is ideal for different physiological procedures, as C29 BRs are biologically less active than C28 BRs.
3. Abscisic Acid
A nonvolatile C15 terpenoid is ABA carboxylic acid, and it was initially presumed that it is made directly from the precursor of the C15 sesquiterpene FDP, or farnesyl diphosphate. Still, trials involving precursors with 18O labels showed that ABA is produced by the C40 carotenoids cleaved from the MEP pathway. Violaxanthin is either converted to 9-
cis-violaxanthin or 9′-
cis-neoxanthin by forming transneoxanthin (C40) [
13,
14]. This process aids in protecting plant tissues from photooxidative stress [
15]. The neoxanthin-deficient Arabidopsis mutant aba4 showed a decrease in ABA production, indicating that neoxanthin is the source of ABA, particularly under water-stress situations. The process of xanthophyll neoxanthin production is still unknown, despite the substantial advancements in our knowledge of the carotenoid biosynthesis pathway. The 2-C-methyl-
d-erythritol 4-phosphate (MEP) route of isopentenyl diphosphate (IPP) synthesis produces carotenoids inside plastids. Plants have virtually figured out the production process for carotenoid pigments. But ABA was not entirely eliminated in the mutant, suggesting that cisviolaxanthin could potentially be used to produce ABA. Furthermore, although lacking 90-cis-neoxanthin, the parasitic plant
Cuscuta reflexa can nevertheless produce ABA [
16]. The cleavage of both xanthins by 9-cisepoxycarotenoid dioxygenase (NCED) to release the C15 compound xanthoxin (C15H22O3) into the cytosol is the last plastidial step in ABA biosynthesis, and this step limits the rate of ABA biosynthesis.
4. Cytokinins
Another class of naturally occurring low-molecular-weight substances, cytokinins (CKs) bind to specific receptors and set off a series of reactions that control plant growth and development. They are mostly recognized by biologists as the chemicals that, along with another hormone called auxin, regulate cell division (cytokinesis). In addition, they play a role in the development of lateral shoots, vascular differentiation, apical dominance, senility and other processes. CKs can have an aromatic side chain or an isoprenoid side chain at the N-6 position of their adenine moiety, as indicated in the introduction. Plant cells can produce isoprenoid CKs through the transfer of a donor C5 isoprenoid unit to an adenine molecule that serves as an acceptor, which can be a tRNA-bound species or a free nucleotide [
17]. DMAPP and HMBPP are the currently recognized viable isoprenoid donors. Both the MEP and MVA pathways in plastids and the cytosol of eukaryotes can produce DMAPP.
Transferring the isoprenoid moiety to adenine, either in its nucleotide forms or coupled to RNA, is the first step in the production of isoprenoid cytokinins. Dimethylallyl pyrophosphate (DMAPP) and (E)-4-hydroxy-3-methyl-but-2-enyl diphosphate are the isoprenoid side chain donors that have been discovered to date. Cytochrome P450 monooxygenase has the ability to further hydroxylate the side chain in the event of DMAPP attachment and isopentenyladenine-type cytokinin production [
18]. Following their eventual liberation from tRNA, the cytokinin nucleotides are hydrolyzed to produce free bases. DMAPP and HMBPP are the currently recognized viable isoprenoid donors. Both the MEP and MVA pathways in plastids and the cytosol of eukaryotes can produce DMAPP.
The MEP pathway active in bacteria and parasites produces only HMBPP [
19]. Adenylate isopentenyl transferases (IPTs) are responsible for catalyzing the N-prenylation of adenine nucleotides. Two types of IPTs are identified: (1) An IPT that functions by adding an isopentenyl unit to a free adenine nucleotide (EC 2.5.1.27) [
20]; and (2) an IPT that performs the same function for tRNA-binding adenine units (EC 2.5.1.8). The isopentenyladenine (iP) nucleotide is the main product of reactions in which DMAPP acts as the IPT’s substrate. On the other hand, the trans-zeatin (tZ) nucleotide is produced by reactions involving the hydroxylated isoprenoid substrate HMBPP (
Figure 1) [
21,
22].
Figure 1. MEP pathway in plants.
5. Strigolactones
Of all the groups, strigolactones (SLs) are newly identified signaling molecules in plants that have hormonal activity [
23]. The primary cause for this discovery was their apparent part in preventing the division of shoots [
24,
25], which seems to be governed by a mechanism encompassing the interaction of auxins, CKs and SLs. Together, SLs also seem to control leaf senility via ethylene, which is gaseous [
26], and to strategically modify overall development to regulate other physiological processes like the distribution of resources in balance. To increase phosphate intake, for example, phosphate scarcity causes plants to uplift SL biosynthesis. This results in modifications to root elongation and the encouragement of fungal symbiosis, indicating that some SLs serve as rhizosphere-signaling molecules [
27]. As per [
28], about 20 SLs are identified and chemically characterized from root exudates of different terrestrial plants. According to [
29], they are tricyclic lactones with three fused rings, A, B and C, along with a butenolide ring (D) joined by an enol ether bond.