Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Pharmacology & Pharmacy
α-亚麻酸 (ALA) 属于 n-3 多不饱和脂肪酸 (n-3 PUFA) 家族,在碳链甲基末端的第三个碳原子上含有碳-碳双键。这个必需脂肪酸家族还包括二十碳五烯酸(EPA)和二十二碳六烯酸(DHA)。ALA因其营养和药用优势而逐渐受到越来越多的关注。研究表明,ALA对包括癌症在内的多种疾病都有有益的作用。
- α-linolenic acid
- anticancer
- cell proliferation
- apoptosis
- inflammatory response
- tumor metastasis
- antioxidant
1. Introduction
Cancer has been a constant threat to human life since its identification, and even when it is treatable, it greatly reduces quality of life. Many studies have shown that ALA exerts significant anticancer effects on multiple cancers [38,39,40,41,42,43,44,45,46,47,48]. In Table 1, a subset of ALA-sensitive cancers is listed, including prostate cancer, BC, hepatocellular carcinoma, colorectal cancer (CRC), and pancreatic cancer. In addition, ALA also exerts effects on many common gastrointestinal tumors and bladder cancer [49,50,51]. As shown in Figure 1, ALA exerts a variety of anticancer effects, including inhibiting proliferation, inducing apoptosis, suppressing tumor metastasis and angiogenesis, and exerting antioxidant effects. To provide a brief introduction to the anticancer effects of ALA, these effects are systematically reviewed, focusing on pharmacological actions and molecular mechanisms.
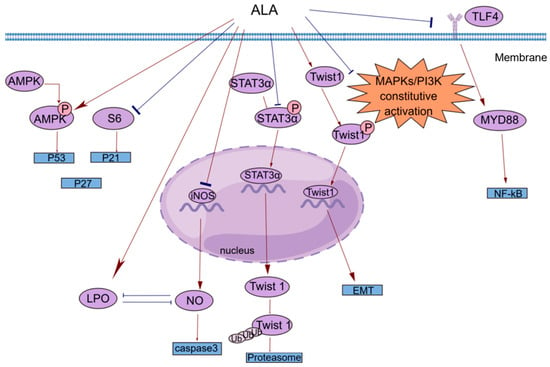
Figure 1. A brief summary of the molecular mechanism of the anticancer effects of ALA. ALA inhibits cell proliferation by regulating the AMPK/S6 axis. ALA can promote cell apoptosis by directly increasing intracellular lipid peroxidation (LPO) or indirectly reducing the accumulation of NO. ALA can suppress tumor metastasis by decreasing the mRNA expression of Twist1 and promoting the degradation of Twist1. The anti-inflammatory effects of ALA may be mediated by blocking the TLR4/MyD88/NF-κB cascade. This figure was constructed with FigDraw (ID: TOUPR3fbb8). There are two kinds of arrows, the flared arrows represent inhibition, and the other is facilitation.
Table 1. The mechanism of the antitumor effects of ALA.
| Cancer | Effect | Effector Molecules | Change in Ex-Pression |
|---|---|---|---|
| PCa (prostate cancer) [52] |
anti-inflammatory effect | PG/LTs | downregulation |
| BC (breast cancer) [38,39] |
anti-inflammatory effect/inhibition of tumor metastasis | COX2/PGE2/Twist 1 | downregulation |
| HCC (hepatocellular carcinoma) [40,41] |
inhibition of proliferation | Farnesoid X receptor | upregulation |
| CRC (colorectal cancer) [42,43] |
induction of apoptosis | caspase 3 | downregulation |
| PCA (pancreatic cancer) [44] |
anti-inflammatory effect | IL-1β/IL-6 | downregulation |
2. Inhibition of Proliferation
Cell proliferation plays a key role in life. Normal cell proliferation is critical for organismal growth, development, tissue repair, and metabolism. However, the abnormal expression of cancer-related genes in cells caused by various factors can lead to uncontrolled cell proliferation, which is an important part of cancer development.
Esophageal tumors can lead to dysphagia and strongly affect patient quality of life. In clinical practice, small and localized tumors are often surgically removed, but there is no way to perform surgery on larger or non-localized tumors. In addition, the resistance of esophageal cancer to chemotherapy has made the need for new therapies more urgent. Hyun-Seuk Moon’s team [53] reported that dietary ALA with or without oleic acid (OA) could inhibit the proliferation of the esophageal cancer cell lines OE19 and OE33 by regulating the AMPK/S6 axis to treat esophageal tumors. OA and ALA promoted the expression of tumor suppressor genes, such as p53, p21, and p27, by activating AMPK and/or decreasing the phosphorylation of S6. This provides a new idea for cancer treatment involving the consumption of ALA-containing foods for therapeutic purposes. Studies have shown that ALA alone or in combination with other drugs can inhibit cell proliferation in a variety of ways. Peroxisome proliferator-activated receptor-γ (PPAR-γ) is a nuclear receptor that regulates lipid homeostasis. As natural ligands of PPAR-γ, fatty acids can inhibit the growth of cancer cells by activating PPAR-γ [54]. Lijun Yang et al. [55] reported that ALA dose-dependently inhibited the proliferation of renal cell carcinoma (RCC) cells by significantly increasing PPAR-γ activity and gene expression and significantly inhibiting cyclooxygenase-2 (COX-2). COX-2 is an inducible enzyme involved in inflammation. Moreover, when ALA was combined with the PPAR-γ agonist rosiglitazone and the COX-2 inhibitor N-(3-pyridyl) indomethacinamide, its inhibitory effect on the proliferation of the human RCC cell line OS-RC-2 was further increased. ALA inhibited the transformation of cervical cancer cells by reducing the expression of the human papillomavirus oncoproteins E6 and E7, restoring the expression of the tumor suppressor proteins p53 and Rb, and reducing the expression of phosphorylated ERK1/2 and p38. Thus, cell proliferation was inhibited [56]. Consequently, it is possible and promising to use ALA in clinical practice to treat associated tumors by preventing tumor cell proliferation.
3. Induction of Apoptosis
Apoptosis is regulated by genes and is a type of programmed cell death. During embryonic development, certain cell populations undergo apoptosis to eliminate certain cells and complete organogenesis. However, mutagenesis by external factors can cause normal cells to become cancerous. The carcinogenesis of normal cells requires uncontrolled cell proliferation, the dysfunction of cell apoptosis, and the dysregulation of apoptotic regulators. In general, cancerous cells escape apoptosis by upregulating antiapoptotic factors and downregulating proapoptotic factors [57].
Most studies investigating the antiapoptotic effects of n-3 PUFAs have focused on major substances such as EPA and DHA, and little research has been conducted on ALA [58,59,60]. However, the functions of these substances are different. For example, one prospective study separated DHA, EPA, and ALA and found that ALA was the only n-3 PUFA that significantly reduced BC risk [17]. Interestingly, this phenomenon of local generalization is not uncommon. Caspases are a class of cysteine proteases that can mediate apoptosis. The apoptotic effect of ALA is closely related to its ability to increase lipid peroxidation [62]. An increase in lipid peroxides may increase the generation of free radicals, and reactive oxygen species (ROS) can directly activate mitochondrial permeability transition, leading to the loss of mitochondrial membrane potential. This results in cytochrome c (cyt c) release and caspase pathway activation. ALA reduced the mRNA expression of inducible nitric oxide synthase (iNOS) to reduce intracellular levels of NO, which can inhibit lipid peroxidation by scavenging free radicals from lipid peroxidation. ALA also inhibited iNOS-induced NO production in a peroxidation-dependent manner, further activating caspase 3 to induce apoptosis [62]. In addition, studies have shown that ALA can promote apoptosis in a variety of ways, such as by upregulating the expression of the proapoptotic gene Bax, downregulating the expression of the antiapoptotic gene Bcl-2, stabilizing hypoxia-inducible factor-1α (HIF-1α) and downregulating fatty acid synthase (FASN) to promote mitochondrial apoptosis [63,64]. This opens up multiple possibilities for the clinical use of ALA.
4. Anti-Inflammatory Response
The inflammatory response is a double-edged sword. When the normal balance of the body is disrupted, immune activity is increased and typically manifests as inflammation. However, the existence of inflammation itself and the changes in the microenvironment caused by inflammation cause certain pathological symptoms. Cancer patients are prone to secondary inflammatory diseases, and cancer may also develop in response to inflammation [65,66]. For example, patients with inflammatory bowel diseases such as ulcerative colitis and Crohn’s disease have an increased risk of CRC and a higher mortality rate than patients with sporadic CRC [67]. The main reason may be the recurrent chronic inflammatory response, which continuously damages the intestinal mucosa. The mucosa is in a state of long-term repair accompanied by intestinal microbial heterotopia and atypical hyperplasia, which can eventually lead to cancer [68]. ALA has been shown to exert powerful anti-inflammatory effects [69], and different epidemiological studies have shown that ALA is inversely correlated with plasma levels of inflammatory factors, including C-reactive protein (CRP), interleukin-6 (IL-6), interleukin-1β (IL-1β), interferon γ (IFN-γ), tumor necrosis factor-α (TNF-α), and E-selectin [25,26]. The potential anti-inflammatory mechanisms of ALA are diverse and include reducing the level of arachidonic acid (AA) in the blood and downregulating COX-2 [70].
A common clinical phenomenon in alcoholic hepatitis patients is the leakage of bacterial endotoxins through the damaged intestinal barrier into the portal vein. The endotoxins then bind to lipopolysaccharide-binding protein (LBP), triggering the TLR4/MyD88/NF-κB inflammatory cascade in the liver. ALA can inhibit the lipopolysaccharide (LPS)-induced inflammatory response by blocking this cascade [71]. N-6 polyunsaturated fatty acids, such as linolenic acid (LA), can be metabolized to AA, which is the precursor of many potent proinflammatory mediators, including prostaglandins (PGs) and leukotrienes (LTs) [72]. ALA intake can reduce blood levels of AA because ALA competes with LA for the enzyme delta-6-desaturase. Moreover, ALA is the preferred substrate of this enzyme. Increasing ALA intake can limit the conversion of LA to AA, thereby reducing the biosynthesis of proinflammatory eicosanoid acid and further exerting anticancer effects [52,73]. COX-2 enzymes are involved in the synthesis of proinflammatory prostaglandins, and one possible mechanism by which ALA inhibits cancer is that it inhibits inflammation through the downregulation of COX-2 [74]. NF-κB plays an important role in the inflammatory and immune responses. COX-2 is a downstream target of NF-κB activation. ALA suppresses tumors by reducing the expression of NF-κB and its target genes in tumor cells [56].
5. Inhibition of Tumor Metastasis
Tumors can spread in vivo to local normal tissues, to nearby lymph nodes, tissues, and organs, or to distant tissues through fluid transport, which is a feature that has made them a leading cause of cancer-related death [75]. Cancer metastasis can be divided into the following steps: tumor growth in situ, angiogenesis, epithelial–mesenchymal transition (EMT), invasion, intravasation, survival in the blood circulation, extravasation, dormancy, and metastatic tumor growth [76].
体内和体外实验表明,ALA在不同程度上抑制了各种癌症的转移。Marianela Vara-Messler等[77]以富含ALA的奇亚籽油和富含LA的玉米油作为对照,给BALB/c小鼠喂食,发现ALA可显著降低小鼠BC的发病率和转移性病变的数量。从机制上讲,ALA可以通过改变细胞膜结构来改变BC细胞的信号传导,从而增加脂肪酸不饱和度并发挥抗癌作用。通过体外实验探索ALA抑制癌症转移的机制,包括EMT和肿瘤血管生成。Twist1 是启动 EMT 和促进肿瘤转移所必需的,并受信号转导和转录 3α 激活因子 (STAT3α) 和丝裂原活化蛋白激酶 (MAPK) 的调节。Shih-Chung Wang等[39]研究了ALA对Twist1和Twist1介导的TNBC细胞迁移的影响,报道ALA可降低Twist1的mRNA表达,减少STAT3α在细胞核中的积累,降低Twist1的蛋白和磷酸化水平。ALA 促进 Twist1 降解,从而消除 EMT 并抑制 TNBC 细胞中 Twist1 介导的迁移。EMT 的标志性特征之一是由于维持上皮细胞接触的粘附连接降解而导致上皮完整性丧失。这种降解的主要驱动因素是基质金属蛋白酶 (MMP) 的蛋白水解消化。在肿瘤中,MMP2 和 MMP9 参与结缔组织降解、肿瘤诱导的血管生成和细胞迁移。ALA抑制肿瘤转移的另一种机制是降低血管内皮生长因子(VEGF)、MMP-2和MMP-9蛋白的表达[56]。作为一种重要的血管扩张剂,NO可以刺激VEGF的产生,并参与VEGF介导的肿瘤血管生成的每一步。NO可以由体内iNOS酶产生。ALA不仅通过降低iNOS的mRNA表达来降低NO水平,而且还增加脂质过氧化。过氧化物的积累增加了自由基的产生;因此,NO的灭活和减少可减少肿瘤血管生成并抑制肿瘤转移[56,62]。
6.抗氧化作用
氧化应激(OS)是指体内ROS产生和消除之间平衡的破坏,主要特征是ROS和活性氮(RNS)等高反应性分子的过度产生。OS与肿瘤的发生发展密切相关[78,79]。Rashmi Deshpande等[62]的研究表明,ALA可以通过刺激ROS产生诱导细胞凋亡来抑制癌症。然而,这项研究是在相对简单的体外实验环境中进行的,体内有更复杂和多样化的机制可以抵消这种影响。Leslie Couedelo等[80]表明,ALA摄入会诱导维生素E的消耗。由于维生素E是一种有效的抗氧化剂,因此可用于捕获产生的自由基而不产生OS。此外,Jin Hyang Song和Teruo Miyazawa报道,在细胞膜中过量掺入n-3 PUFAs会增强细胞膜对脂质过氧化(lipid peroxidation, LPO)的敏感性并诱导OS,从而产生不良影响,但这些研究是基于高脂肪饮食[81]。总之,当ALA添加到饮食中时,应考虑其对整个身体的影响,也应考虑摄入量。
亚麻籽油(FO)富含ALA,常被用作治疗癌症和改善健康的膳食补充剂。Jyoti Sharma等[82]研究了由7,12-二甲基苯并-[a]烷(DMBA)与巴豆油联合诱导的皮肤癌小鼠,发现FO通过增加皮肤和肝脏中酶和非酶抗氧化剂的水平来清除自由基,包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GPx)和谷胱甘肽(GSH)。SOD、CAT 和 GPx 是重要的抗氧化酶,它们协同工作以防止细胞内 ROS 水平过高。SOD的主要作用是加速超氧阴离子向过氧化氢的歧化,然后CAT和GPx将产生的过氧化氢转化为无害物质[83]。GSH的自由基清除作用是由其结构中的活性巯基-SH基团介导的,这些基团容易通过氧化脱氢。此外,GSH可以与GPx结合,需要不同的二级酶和辅因子,包括烟酰胺-腺嘌呤二核苷酸磷酸(NADPH)才能发挥其作用[82]。NADPH氧化酶是脉管系统中ROS的主要来源,由催化亚基和调节性胞质亚基组成。Hao Han[84]对动脉粥样硬化进行了研究,发现FO可以通过降低NADPH氧化酶催化亚基的mRNA和蛋白表达来抑制NADPH氧化酶,从而调节细胞质亚基的表达,降低丙二醛水平,增加GSH水平,发挥抗氧化作用。富含ALA的FO主要作为食用油食用,这种营养素与肠道微生物群之间无疑有着密切的联系。Xiaoyan Sun等[85]采用FO饮食研究了老年大鼠结肠上皮细胞的抗氧化能力,结果表明,FO给药后老年大鼠的抗氧化水平显著增加。从机制上讲,FO的摄入可能改变了一些肠道细菌。OS可以在肿瘤发展的整个过程中发挥作用,在肿瘤发展的不同阶段服用ALA可以减缓对身体的毒性作用。
This entry is adapted from the peer-reviewed paper 10.3390/jpm14030260
This entry is offline, you can click here to edit this entry!