1. Introduction
The corneal epithelium is the outermost part of the cornea and consists of three cellular layers: the superficial layer, middle wing layer, and the innermost basal cell layer with Bowman’s membrane in humans separating it from the corneal stroma [
1]. It plays the role of a protective barrier as well as a refractive structure due to its avascular character. Its homeostasis is maintained through a complex regenerative process which takes about 10 days and involves proliferation and migration of epithelial cells [
2]. In addition, the lacrimal gland function, tear film, and corneal nerves are pivotal in maintaining the health and integrity of the corneal epithelium [
3].
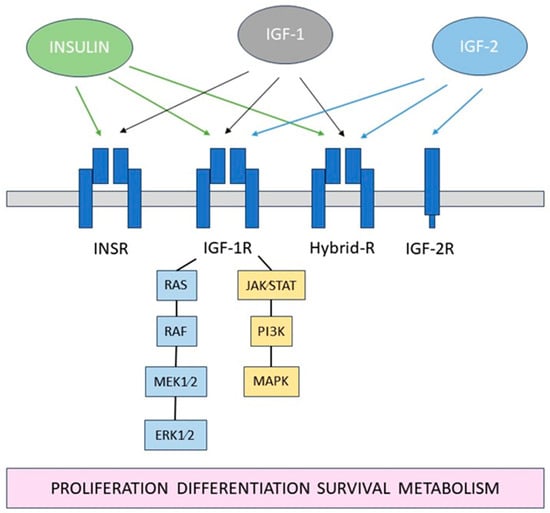
The insulin-like growth factor (IGF) system is a complex network of hormones and proteins that play crucial roles in cell growth, development, and metabolism [
19]. It consists of insulin, insulin-like growth factor 1 and -2 (IGF-1, IGF-2), their receptors: insulin receptor (INSR), IGF type 1 and 2 receptors (IGF-1R, IGF-2R), as well as several IGF-binding proteins (IGFBPs),
Figure 1.
Figure 1. Schematic diagram of the IGF system.
2. The Role of Insulin and Effect of Diabetes
Insulin is a polypeptide hormone consisting of two A and two B chains, produced by pancreatic beta cells and secreted in response to a high blood glucose level [
19]. It has metabolic effects and plays a role in various phases of the cell cycle, from growth to apoptosis. The presence of INSR in the cornea was first demonstrated by Naeser [
41]. Alternative splicing of INSR occurs at exon 11, leading to the generation of two distinct isoforms: INSRA and INSRB [
19]. An immunohistochemical analysis by Rocha et al. demonstrated that INSR is expressed in the corneal epithelium [
42]. In their study of human corneas, INSR was predominantly localized within the cytoplasm and plasma membrane in the wing and superficial cell layers, with noticeable variability in its expression across the basal and intermediate suprabasal cells (
Table 1).
Table 1. Summary of the IGF system receptors and their localization in the corneal epithelium.
| Receptor |
Localization |
Ref. |
| Insulin receptor |
Plasma membrane and cytoplasm; mainly in the wing and superficial cell layers |
[42] |
| |
Nucleus |
[43] |
| |
Mitochondria |
[22] |
| IGF-1R |
All layers of the cornea; mainly around cellular nuclei of actively differentiating epithelial cells |
[43] |
| |
Plasma membrane and cytoplasm |
[42] |
| |
Mitochondria |
[22] |
| Hybrid- R |
Plasma membrane and nucleus |
[44] |
| IGF-2R |
Central and peripheral epithelium with higher expression in the periphery following corneal injury |
[34] |
| |
Primarily in the basal corneal epithelium in murine and porcine corneas |
[35] |
Diabetes mellitus, a chronic metabolic disorder characterized by hyperglycemia, results from a deficiency in insulin secretion, impaired insulin action, or a combination of both [
45]. The two primary forms of diabetes, type 1 and type 2, differ in their pathophysiology but share the common feature of dysregulated glucose metabolism. In type 1 diabetes, an autoimmune response leads to the destruction of pancreatic beta cells, resulting in insufficient insulin production, while type 2 diabetes involves a combination of insulin resistance and relative insulin deficiency. Diabetes can significantly impact the eye, including the ocular surface, leading to dysfunction of epithelium and development of diabetic keratopathy.
Unlike most tissues, where insulin stimulates glucose uptake through the glucose transporter-4 (GLUT4), corneal epithelium is insulin-independent [
46]. Glucose uptake in epithelial cells occurs through constitutively active glucose transporters, GLUT1, which undergo upregulation in case of a high metabolic demand, such as wound healing, similarly in diabetic and non-diabetic corneas.
Culture studies with human corneal epithelial cells demonstrated that insulin induces phosphorylation of extracellular signal regulated kinase (ERK 1/2), PI3-kinase [
20] and epidermal growth factor receptor (EGFR), thereby promoting cell migration and wound healing [
21]. Interestingly, increased expression of PI3K pathway kinases occurs in canine corneal cells following insulin treatment, contrasting with the observed lack of analogous effects in vitro in human corneal cells [
47]. Within the corneal epithelium, insulin influences PTEN-induced kinase 1 (PINK-1)-mediated mitophagy and the mitochondrial accumulation of insulin receptor (INSR). Interactions between INSR and the voltage-dependent anion channel-1 (VDAC1) prevent fragmentation and altered polarization of mitochondria, as well as facilitate PINK-1-mediated mitophagy [
22]. In diabetic rats, the process of histone H3 acetylation is reduced in corneal epithelial cells, resulting in compacted chromatin organization in nuclei characterized by increased size and elevated DNA ploidy [
48].
A study by Song et al. showed that insulin can normalize the circadian rhythm of corneal cell mitosis via five main clock genes (Clock, Bmal1, Per2, Cry1, and Rev-erbα) whose expression is affected in diabetes [
23]. Moreover, innate-like lymphocytes, such as γ δ T-cells expressing chemotactic factor IL-17 for neutrophils and monocytes, were found to be recruited to the corneal limbus in a diurnal pattern. In diabetes, the limbal cell migration is increased, potentially leading to the inflammatory state delaying wound healing, but is restored upon systemic insulin administration.
Insulin is secreted from the lacrimal gland and present in the tear film at a mean concentration of 0.404 ± 0.129 ng/mL, which is reduced in fasted individuals and shows no difference related to gender [
42]. In diabetes, its secretion is reduced due to damage to the lacrimal gland and reduced corneal sensation, caused by hyperglycemia and oxidative stress [
49]. In rat models of diabetes, histological analysis showed an increased lipofuscin level and higher malonaldehyde as well as peroxidase activity in the lacrimal glands compared to healthy animals [
50]. Insulin signaling in rat lacrimal glands becomes impaired in the fourth week of diabetes, with the lacrimal gland serving as an extra pancreatic source of insulin for at least 4–7 weeks [
51,
52].
Several studies reported ocular surface abnormalities in patients with diabetes, such as reduced tear breakup time, lower tears secretion, and increased level of inflammatory markers: NPY, STAT-5 ICAM-1, and TNF-α [
53,
54,
55]. Clinically, diabetic ocular surface complications include reduced corneal sensitivity and delayed epithelialization leading to dry eye syndrome, punctate corneal epitheliopathy, recurrent erosions, persistent epithelial defects, and neurotrophic keratopathy [
49,
56]. On a histological level, reduction in basal epithelial cell density and size, and increased intercellular space as well as increased corneal epithelial basement membrane thickness and irregularity were demonstrated in diabetic corneas [
57,
58,
59,
60,
61]. It is postulated that diabetic ocular changes might be in part explained by the dysregulation of a pathway involving the opioid growth factor (OGF), i.e., [Met5]- enkephalin binding to nuclear-associated receptor (OGFr) [
62]. In diabetes, serum OGF levels are elevated, and insulin may affect the OGF-OGFr axis [
56,
63,
64].
A fundamentally negative impact of diabetes on the corneal epithelium is closely linked to its effect on LESC functioning [
24]. A significant reduction in Palisades of Vogt in all four limbus quadrants was demonstrated in patients with type 2 diabetes using in vivo confocal microscopy [
65]. In the same study, a higher risk of stem cell damage was noted in those with a high-density lipoprotein, triglycerides, and total cholesterol level above 1.215 mmol/L, 1.59 mmol/L, and 4.75 mmol/L, respectively.
Immunohistochemistry analysis of corneas from diabetic patients revealed a reduction in putative limbal stem cell markers, including ATP-binding cassette superfamily G member 2 protein (ABCG2), N-cadherin, ΔNp63α, K15, K17, K19, and β1 integrin [
28]. This decrease in marker expression was associated with lower immunoreactivity and a diminished number of detected cells, indicating potential depletion or dysfunction of LESCs. Moreover, the ex vivo diabetic limbus was characterized by irregular epithelial basement membrane and reduced expression of laminin γ3 and fibronectin.
In another study, a reduction in expression of putative stem cell markers K15 and ΔNp63α was demonstrated in limbal epithelial stem cell (LESC)-enriched cultures obtained from the corneoscleral rims in diabetic patients with changes reversed by adenoviral gene therapy [
27]. In an animal model of type 2 diabetes, reduced expression of corneal stem/progenitor cell markers, including Hes1, Keratin15, and p75, was observed in mice corneas [
25]. Similarly, in mice with type 1 diabetes, the expression of putative LESC markers K15, ∆Np63α, and glycoprotein hormone alpha-2 (GPHA2) was reduced in the limbus [
26].
A study employing a new method of objective quantification of immunofluorescence, aiming to overcome limitations of manual grading, observed a reduction in the putative LESC marker K14 in the limbus of diabetic mice [
24]. Furthermore, reduced expression of putative limbal epithelial stem cell markers, such as paired box protein-6 (PAX6), ∆Np63α, K15, K17, and membrane transporter ABCG2, was demonstrated in cultured diabetic human limbal epithelial cells, which was associated with slower corneal epithelial wound healing [
29]. It is crucial to note, however, that several cell markers for LESCs can also be identified in other parts of the eye, and their use has limitations in discriminating pure stem cells from early transient amplifying cells [
24,
66].
The nervous system plays an instrumental role in maintaining the homeostasis of the corneal epithelium. Nerve branches of the sub-basal corneal nerve plexus are considered an important source of substances contributing to ocular surface health. Importantly, corneal innervation impacts a stem cells niche [
67], with a limbal stem cells reduction reported following the destruction of the ophthalmic branch of the trigeminal nerve [
25].
Detrimental effects of diabetes on corneal sensitivity, nerve fiber length, and density in both humans [
68,
69,
70,
71,
72] and animals [
25,
73,
74,
75,
76,
77,
78] have been reported in numerous studies and reviewed elsewhere [
79,
80,
81]. Recently, a 3D tissue model of the human cornea was employed to demonstrate the degenerative effects of hyperglycemia on corneal nerves [
82]. On a cellular level, pannexin1 channels present in corneal synaptosomes were found to be more glycosylated, characterized by enhanced membrane localization and leading to increased ATP release in diabetic subjects compared to non-diabetic controls [
83]. Animal studies showed that insulin stimulates corneal nerve regeneration and expression of a limbal stem cell marker (DNp63) via Wnt signaling [
84], and when applied topically, it exerts neuroprotective properties in diabetic rats [
76]. In patients with type 2 diabetes, a nerve regenerative process can be limited by insulin resistance [
85]. Interestingly, a prediabetic state is characterized by increased parameters of intra-epithelial corneal basal nerves, which could be attributed to the neurotrophic effect of higher insulin levels [
78].
MicroRNAs (miRNAs) are 18–25 nucleotides long non-coding RNAs that downregulate the expression of genes at a post-transcription level by binding complementary mRNAs [
88]. They are involved in numerous cellular processes and can modulate multiple genes, making them a valuable research focus in regenerative medicine [
89]. Studies suggest that they also play a role in LESC-associated processes, such as macropinocytosis, autophagy, and the expression of putative stem cell markers [
90,
91]. Funari at al. reported on the dysregulation of miRNA expression in human autopsy diabetic corneas, which was associated with abnormal wound healing [
92]. Microarray analysis demonstrated that among the 29 miRNA studied, miR-146a, 21, and 424 were the most upregulated, while miR-509-3p and 143 were expressed at the lowest level. This is in line with other reports, which indicate that overexpression of miR-146a in the diabetic limbus may result in a reduced corneal inflammatory and healing response [
24,
93,
94]. In another study, genome-wide sequencing was applied, identifying differences in expression profiles of 20 miRNA between normal and diabetic human corneas [
95]. Results showed that miR-10b was upregulated in the diabetic limbus, with a higher increase observed in type 1 compared to type 2 diabetes. Moreover, altered expression of miRNA in exosomes have been demonstrated in diabetic limbal stem cells [
96].
Overall, diabetes may affect the function of LESCs through various mechanisms and can potentially lead to severe, persistent corneal erosions. However, despite the detrimental effect on the cornea, the limbal barrier tends to be preserved, with no development of conjunctivalization or neovascularization characteristic for limbal stem cell deficiency [
24].
Animal studies have demonstrated the beneficial effect of insulin treatment on epithelial wound healing in diabetic animals [
97,
98]. Klocek et al. found that corneal abrasions in type 1 diabetic mice, treated with topical insulin, reduced in size by 29% compared to the controls after 16 h [
97]. Moreover, Zagon et al. reported a comparable rate of corneal healing in diabetic rats with an insulin implant and normal animals [
98].
The first therapeutic use of topical insulin in humans was described by Aynsely in 1945 in a case series of patients with corneal ulcers [
99]. Successful treatment with topical insulin for corneal conditions involving the epithelium has also been reported in subsequent studies summarized in
Table 2.
Table 2. Published studies evaluating results of a treatment of corneal conditions with epithelial defects with a topical insulin.
| Ref. |
Study Design |
Diagnosis |
No. of Eyes |
Mean Age
(Years) |
Eyes with Complete Epithelialization (%) |
Mean Time to Epithelialization
(Days) |
| [100] |
Randomized controlled trial |
Postoperative corneal epithelial defect after vitreoretinal surgery in diabetics |
A—8
B—8
C—8 |
A—62.62 ± 5.99
B—56.12 ± 7.77
C—55.75 ± 6.64 |
A—100
B—100
C—100 |
All eyes healed within 6 days
100% eyes in A, 75% eyes in B and 62.5% eyes in C group healed within 3 days. |
| [101] |
Randomized clinical trial |
Postoperative corneal epithelial defects after vitreoretinal surgery |
19 |
57.05 ± 12.33 |
100 |
3 |
| [102] |
Prospective interventional, single-center case series |
Refractory persistent epithelial defects |
11 |
45.4 ± 25 |
82 |
62.3 ± 34.6 |
| [103] |
Prospective non-randomized hospital-based study |
Refractory persistent epithelial defects |
21 |
72.2 |
81 |
34.8 ± 29.9 |
| [104] |
Prospective non-randomized hospital-based study |
Recurrent epithelial erosions |
15 |
29.00 ± 8.72 |
100 |
21 |
| [105] |
Retrospective, observational |
Refractory neurotrophic keratopathy (NK) in stages 2 and 3 |
21 |
61 |
90 |
18 ± 9 in NK stage 2; 29 ± 11 in NK stage 3 |
| [106] |
Retrospective, consecutive case–control series |
Refractory persistent epithelial defects |
61 |
71.5 ± 19.3 |
84 |
32.6 ± 28.3 |
| [107] |
Retrospective
case series |
Refractory neurotrophic corneal ulcers |
6 |
36.5 |
100 |
7 to 25 |
| [108] |
Retrospective
Case series |
Corneal epithelial erosions induced during vitreoretinal surgery in diabetics |
5 |
49 |
100 |
2.5 ± 0.6 |
| [109] |
Retrospective case series |
Dry eye disease |
32 |
61.3 ± 16.8 |
- |
- |
| [110] |
Case report |
Corneal ulcer following chemical injury |
1 |
41 |
100 |
60 |
| [111] |
Case report |
Bilateral Neurotrophic keratitis |
2 |
55 |
100 |
7 |
| [112] |
Case report |
Neurotrophic keratopathy after resection of acoustic neuroma |
1 |
45 |
100 |
14 |
| [113] |
Case report |
Neurotrophic keratopathy |
1 |
40 |
100 |
20 |
| [114] |
Case report |
Neurotrophic keratopathy |
1 |
64 |
100 |
30 |
Interestingly, a recent culture study with ocular surface cells showed that drops from plasma rich in growth factors were superior to topical insulin at two different concentrations (1 and 0.2 IU/m) in promoting wound healing and reducing the fibrosis process [
115]. Currently, no universal protocol exists regarding treatment with topical insulin, as studies apply different preparation methods, dosages, concentrations, and types of drops [
116,
117].
Several studies have demonstrated novel ways of delivering insulin to the eye. A new delivery system, containing chitosan/poloxamer gel loaded with chitosan microparticles, has been reported to increase the local bioavailability of topical insulin in the treatment of the ocular surface in diabetic rats [
118]. Additionally, electrospun fiber mats were shown to be effective in delivering insulin to the porcine cornea [
119]. Recently, a convolutional neural network statistical analysis was applied to demonstrate the therapeutic effect of insulin liposomes on corneal epithelial defects in rats [
120]. In a study of alkali-burned corneal models, a nano-system combining liposomes and trimethyl chitosan was used to deliver insulin and vascular endothelial growth factor small interfering RNA [
121]. This combined therapy proved effective in reducing oxidative stress, increasing epithelialization, and inhibiting corneal neovascularization. Metabolomic analysis demonstrated that the therapeutic effect was possibly linked to insulin inhibiting the ferroptosis signaling pathway.
This entry is adapted from the peer-reviewed paper 10.3390/biology13030144