2. Metalloproteinases and the Kidney
MMPs are classified, according to their structure (or function) and the substrate selectivity, into six groups: Collagenases (MMP-1, MMP-8 and MMP-13), which cleave native collagen and with possible antifibrotic function; Gelatinases (MMP-2 and MMP-9), whose function is to cleave denatured collagens, type IV collagens in basement membranes and some chemokines; Stromelysins (MMP-3, MMP-10, MMP-11 and MMP-19), which degrade a number of substances such as fibronectin, laminin but are unable to cleave native collagen; Matrilysins (MMP-7 and MMP-26) act by degrading ECM components (laminin and entactin); Membrane-type MMPs (MMP-14, -15, -16, -17, -24, and -25), so called because they are anchored to the exterior of the cell membrane; and other MMPs that are tissue or cell-type specific [
10,
13]. The MMP activity is modulated by a series of four known enzymes called tissue inhibitors of metalloproteinases (TIMPs). TIMPs participate either in the activation or inhibition of MMP activity and, like MMPs, regulate several cellular functions such as cell proliferation, apoptosis, and angiogenesis [
14]. It has been demonstrated that MMPs exert a role in the development of proteinuric kidney diseases in humans. Indeed, a number of studies depicted increased serum and urine levels of MMP-2, -8, and -9 in diabetic patients, with MMP-9 in particular being positively correlated with the degree of proteinuria in these patients [
15,
16,
17,
18]. It is remarkable that, in addition to MMP-9, urinary levels of neutrophil gelatinase-associated lipocalin (NGAL) have been found to increase in patients with diabetic nephropathy [
19]. NGAL and MMP-9 are coexpressed, and their interaction prevents the degradation of MMP-9. It has, thus, been postulated that the increase in NGAL may prolong the action of MMP-9 as a trigger of kidney damage [
20]. MMP-7, which is normally expressed in the proximal and distal convoluted tubules, as well as in the collecting duct, is found to be overexpressed in diabetic patients where it is also inversely correlated with the degree of kidney function [
21]. Other than in diabetic patients, MMP expression is altered in many other glomerular diseases. Typical patterns of MMP-2 and MMP-9 are differentially expressed in patients with focal segmental glomerulosclerosis, minimal change disease, membranous nephropathy, and ANCA-associated vasculitis [
22,
23,
24,
25,
26]. Regardless of the specific kidney disease involved, several mechanisms of damage have been put forward to explain the pathophysiologic effects of MMPs (
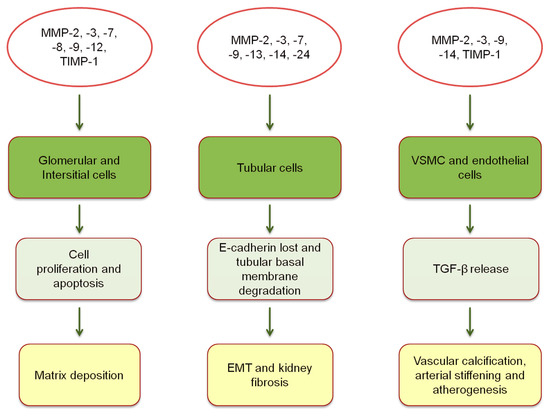
Figure 1).
Figure 1. Expression of MMPs and TIMPs, and pathophysiological mechanisms of vascular and kidney damage [
10,
11,
27,
28,
29,
30,
31]. EMT, epithelial-mesenchymal transition; VSMC, vascular smooth muscle cells; TGF-β, transforming growth factor-β.
MMPs intervene in all phases of renal fibrosis, from infiltration of mononuclear cells to cells proliferation and scarring. All these processes lead to a progressive decline of renal function in CKD patients. Henger and Colleagues performed a hierarchical clustering analysis to assess the differential gene expression in human kidney fibrosis. Interestingly, they observed that several MMP (MMP-3, -13, -14) genes were upregulated in different degrees of fibrosis [
32]. With respect to inflammation, different MMPs play different, often opposing, actions. MMP-7 and MMP-9 expand inflammatory processes, particularly by their chemotactic effect on human dendritic cells [
33,
34]. Conversely, MMP-13 and MMP-14 have been hypothesized to act as anti-inflammatory mediators [
35]. MMP-3 may also promote epithelial-to-mesenchymal transition, the conversion of tissue phenotype, from the epithelial to fibroblastic, thereby accelerating fibrosis [
36]. Abnormalities in the accumulation/degradation of ECM due to imbalanced levels of MMPs and TIMPs have also been described in rat and humans [
37,
38,
39]. Indeed, downregulation of MMP-1 and the overexpression of TIMP-1, MMP-2, MMP-7, and MMP-9 are associated with a profibrotic effect, as well as with a destructive effect on renal parenchyma [
38,
39]. Moreover, increased TIMP-1 plasma levels were predictors of incident CKD, regardless of other systemic and inflammatory biomarkers (C-Reactive Protein or Brain Natriuretic Peptide) and many clinical parameters (liver function, concomitant lipid-lowering, or antihypertensive medications) [
40].
3. Vascular Effects of Metalloproteinases in CKD Patients
The structural remodeling of ECM, together with the profibrotic effect of MMPs, are detrimental for other organs apart from the kidney. Elevated blood concentrations of TIMP-1 have been associated with an increased risk of developing chronic heart failure (CHF) and, in patients already diagnosed with CHF, they were predictors of poor prognoses [
41,
42]. Moreover, the increase in MMP-9 and TIMP-1 conferred risk of all-cause mortality and incident CV disease in community studies [
43,
44]. Possible explanations for the relationship between MMPs and CV risk are varied. Whereas all MMPs are likely risk factors for atherosclerosis and cardiac dysfunction, a more specific mechanism of damage has been postulated for MMP-9 and TIMP-1. Indeed, MMP-9 is involved in the intracellular cleavage of myosin filaments, a mechanism that leads to ventricular hypertrophy [
45]. TIMP-1 has shown a direct relation with the left ventricular mass in the Framingham Heart Study participants [
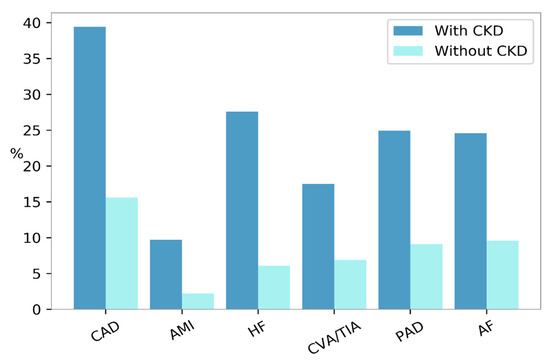
41]. The CKD condition is associated with an increased prevalence of CV morbidity and mortality. The United States Renal Data System (USRDS) showed that the frequency of each CVD, including myocardial infarction, coronary artery disease, and peripheral vascular disease was higher among patients with CKD compared with those without (
Figure 2) [
9].
Figure 2. Prevalence of cardiovascular diseases according to the presence (dark blue bars) or absence (turquoise bars) of Chronic Kidney Disease (CKD) in the United States, in the year 2015. AF, atrial fibrillation; AMI, acute myocardial infarction; CAD, coronary artery disease; CVA/TIA, cerebrovascular accident/transient ischemic attack; HF, heart failure; PAD, peripheral arterial disease.
The CV burden in these patients is significant if one considers that the rate of CV events is similar to that of reaching ESKD [
46,
47]. As a result of this epidemiological and public health evidence, great effort has been placed in finding early atherosclerosis biomarkers that predict CV events in CKD patients and serve as a possible target for new therapeutic agents [
48]. MMPs were included in the set of investigated substances. Indeed, MMP-2 was positively associated with carotid Intima-Media Thickness (cIMT) and abdominal aortic calcification, suggesting that an association between this MMP and subclinical atherosclerosis is plausible [
49,
50,
51]. Moreover, MMP-2 was also found to be higher in patients with a positive history of CV disease vs no history of CV disease [
52,
53]. MMP-9 was also strongly associated with cIMT, the development of carotid plaques, and systemic atherosclerosis [
49,
50]. MMP deregulation is intensified in patients with advanced CKD stages that are also associated with a reduction of their clearance. All these mechanisms enhance the inflammatory process that is chronically activated in CKD patients, due to oxidative stress, the uremic milieu, and the metabolic acidosis [
54]. The combination of uremic status and imbalance in pro-inflammatory substances (such as MMPs) accelerates the atherosclerotic process, arterial stiffness, and vascular calcification, and impairs the vascular repair process as well [
55]. Circulating MMP-2, -9, and -10 have been found to increase in CKD patients, and have been implicated in the vascular damage process. Moreover, MMP-2 and -9 are able to reduce the plaque stability in advanced CKD stages, thus rendering the plaque itself more prone to rupture [
56,
57].
4. Metalloproteinases and Peripheral Vascular Disease
MMPs play a role in the pathogenesis and prognosis of arterial and venous disease. With respect to the damage occurring in the arterial tissue, multiple studies have shown that MMPs are involved in one or more steps of atherogenesis and aneurysm development [
58]. Evidence that confirms this hypothesis comes from both basic and clinical studies. During the progression of atherosclerotic plaques, a number of MMPs are produced, including TIMP-1, MMP-1, -2, -3, -9, and -14 [
59]. An in vivo study on Fischer male rats showed that TIMP-1 directly regulates the smooth muscle cell migration [
27]. A similar function has been recognized in human studies on MMP-14 and TIMP-1, with being both involved in the process of cellular migration to the plaque fibrous cap and plaque inflammation [
28,
29]. MMP-9 also contributes to the destruction of the fibrous cap itself in patients with increased CV risk [
30]. Interestingly, increased concentrations of MMP-2, -3, and -9 have been found within the aneurysm human tissue, being mainly produced by the macrophages localized in the aneurysm wall [
31]. These different enzymes are differentially expressed according to the aneurysm dimensions and severity [
60,
61]. MMP-2 is increased in small aneurysms (<5.5 cm), whereas MMP-9 is dominant in large aneurysms (5.5–7 cm). Moreover, it has also been shown that different localizations of MMP activity within the aneurysm wall modify the risk of rupture [
60]. In clinical studies, abnormal circulating levels of MMP-1, -2, -8, -9, and TIMP-1 were found in patients with Peripheral Arterial Disease; their increase was attributed to the presence of ischemic tissue [
12,
62,
63]. Circulating levels of MMP-1 and -8 were also found as predictors of poor prognosis, in terms of major amputation or death, in patients who underwent lower extremity bypass [
64]. These multiple findings enhanced the importance of MMPs as biomarkers of arterial disease severity that also provide important prognostic information in clinical practice.
Regarding venous diseases, it has been observed that alterations in ECM remodeling are common in the case of varicose veins and chronic venous insufficiency (CVI). With regards to varicose veins, the expression of MMP-1, -2, -3, -7, and -9 is increased, particularly in the smooth muscular cell of the vein wall, both in human and mice models [
65,
66]. Moreover, an analysis of human saphenous vein showed that this expression is even higher in varicose veins with inflammation, as compared to those without inflammation [
65]. Mechanisms underlying the association between MMPs and varicose vein physiopathology might involve the effect of MMPs on ECM degradation and the relaxation of the venous wall [
66]. An upregulation of MMP-1, -2, -9, and -13, together with a downregulation of TIMP-1 and TIMP-2, have been described in patients with CVI [
67,
68]. The distribution of MMP varies based upon the stage (from CVI to active wound) and cells, suggesting that MMP-1 and TIMP-1 are needed in the re-epithelialization phase, while MMP-9 and -13 primarily participate in the remodeling of the collagenous matrix [
69].
5. Synthetic Metalloproteinases Inhibitors in Experimental and Clinical Research
MMP activity is regulated at different levels, including either intracellular (mRNA expression and post-translational modification of MMP structure) or extracellular (stimulation or inhibition of their enzymatic activity from endogenous or exogenous substrates) process. The net activity of MMPs is a crucial step, since its up- or down- regulation could affect MMP activity, and ultimately lead to metabolic diseases, cancer, cardiovascular, and renal disorders [
72]. For these reasons, new pharmacological agents that interfere with MMP activity have been developed and utilized as potential tools that could benefit a wide spectrum of patients. Synthetic MMPs inhibitors (MMPs-I) include broad-spectrum and specific MMPs-I. The vast majority of these compounds contain Zn
2+ in their structure, and have structured as Zn
2+ binding globulin (ZBG). Indeed, ZBGs inactivate MMPs by displacing the Zn
2+- bound water in the MMPs, and favor the anchorage of the drug to the MMPs substrate binding-pocket [
73]. ZBGs encompass hydroxamic acids Batimastat (BB-94), Marimastat (BB-2516), and Ilomastat (GM6001), that displays a broad-spectrum inhibition of MMPs. More selective ZBGs molecules have also been developed, and include hydrazides and sulfonylhydrazides that specifically inhibit MMP-1, -2, and -9. Hydroxamic ZBGs are effective, but they have poor oral bioavailability and, by inhibiting multiple MMPs, cause several musculoskeletal side effects [
74]. Hence, heterocyclic bidentate chelators have been developed that have shown more biostability and lower toxicity in cells assays. Tetracyclines are antibiotic molecules that, by chelating Zn
2+ ion, are able to inhibit MMPs. Doxycycline inhibits MMP-2 and-9 [
75]. Chemically-modified tetracyclines reach higher plasma levels for prolonged periods of time, require less frequent administration, are associated with lower rate of side effects when administered orally, and are thus preferred over conventional tetracyclines [
76]. Apart from zinc-based compounds, several MMPs-I act by a noncompetitive, nonzinc-binding, mechanism of inhibition. They show high selectivity that minimizes the side effects, and thus, are considered very promising molecules [
77]. MMPs-Is have already been used in pilot studies in mice and in human models of kidney damage [
11]. MMPs-I BB-1101 has shown to reduce proteinuria in rats with anti-Thy1.1 nephritis, an experimental model of glomerular damage induced by antibody against Thy 1 gene [
78]. A similar effect on proteinuria was found with the MMPs-I BB-94 in an experimental model of kidney allograft rejection in mice and the tetracycline antibiotic doxycycline in human patients with diabetic nephropathy already under renin-angiotensin-aldosterone inhibition [
79,
80]. Interestingly, doxycycline also reduced the aneurysm expansion in small randomized clinical trials enrolling patients with abdominal aortic aneurysm [
81,
82].