Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Anatomy & Morphology
Breast Cancer (BC) is one of the most common cancer types worldwide, and it is characterized by a complex etiopathogenesis, resulting in an equally complex classification of subtypes. MicroRNA (miRNA or miR) are small non-coding RNA molecules that have an essential role in gene expression and are significantly linked to tumor development and angiogenesis in different types of cancer. miR-125 is a highly conserved family of microRNAs whose members have also been found in nematodes (named lin-4 in 1993, the first miR described ever).
- breast cancer (BC)
- MicroRNA (miR)
- non-coding RNA
- competing endogenous RNA (ceRNA)
1. microRNA Nomenclature
MicroRNAs are short (20–25 nucleotides), single-stranded, non-coding RNA molecules whose main function is gene expression control, mainly silencing. They exert this downregulation by binding the 3′ end of target mRNA(s) through sequence homology and promoting either their degradation or impairing their translation [59]. Over 2500 miR have been estimated to be encoded in the human genome, regulating over 60% of human genes [60]. In addition, thanks to imperfect sequence pairing, they can also bind multiple targets, thus amplifying their intracellular action.
Beyond the number identifier (ID, usually higher for miR described chronologically later), additional nomenclature rules are established to identify unequivocally each miR [61,62]. miRs with almost identical sequence are identified by a progressive lowercase letter after the identification number (miR-XXXa, miR-XXXb, etc.), while miRs with identical sequence but mapping to different genomic locations are indicated by a progressive number separated from the ID number by a dash (e.g., miR-XXX-1, miR-XXX-2, etc.). To further distinguish molecules of different species, an additional three-letter code and a dash may be added at the beginning of the miR name (i.e., hsa-miR-XXX indicates a human–Homo sapiens–miR). Finally, the mature, single-stranded miR can be obtained either from the 5′ end or the 3′ end of its double-stranded miR precursor (pre-miR). Notably, sometimes both strands can—separately—be used for mRNA regulation, resulting in 5p and 3p miR forms if both are present and functional in the cell and the two miR are roughly equivalent in their intracellular amount; instead, if both are present but one is significantly more abundant than the other, then the rarer one has an asterisk at the end of its name (e.g., miR-XXX-5p*).
2. Role of miR in BC
Increasing evidence shows that miRs represent a central hub of gene expression control in human carcinogenesis, and, from this perspective, BC is not an exception [63,64].
Significantly, miR-21 has been shown to be responsible for the development of multidrug resistance [65], and it modulates the resistance of BC cells to doxorubicin by targeting PTEN [66]. In addition, miR-21 also plays a central role in BC proliferation and metastasis by targeting LZTFL1 [67]. Additional miR-21 targets involved in cell proliferation, metastasis, epithelial-to-mesenchymal transition (EMT), and apoptosis in BC include IGFBP3, TPM1, PCD4, and TGF-beta1 [68].
Another player in BC etiopathogenesis is miR-106a, which promotes cancer progression through the downregulation of RAF-1 [69], P53, BAX, and RUNX3 and the upregulation of Bcl-2 and ABCG2; it also confers cisplatin resistance upon its upregulation [70,71].
Upregulation of miR-155 causes telomere fragility through its action on TRF1, a component of the shelterin complex [72]. Interestingly, this miR, together with miR-10b, miR-34a and miR-141, is also a possible candidate for building a panel of circulating miR useful for non-invasive detection of this tumor [73].
Conversely, downregulated miR-141 has been reported to be a typical feature of BC, where its target is ANP32E [74], which, in turn, induces tumorigenesis in triple-negative BC (TNBC) cells by upregulating E2F1 [75]. Instead, high miR-141-3p expression is typical of grade III BC compared to grade II and, together with miR-181b1-5p and miR-23b-3p, it is a useful marker not only to discriminate between malignant and benign breast tissues but might also help in distinguishing TNBC from other molecular subtypes of BC [76].
The let-7 family of miR are tumor suppressors in several cancers, including BC, and their members can also be detected as circulating biomarkers [77]. Let-7 action involves the control of ERCC6 expression [78], and its overexpression could inhibit BC cell proliferation.
Another circulating marker of BC is miR-335 [79], which exerts its effects by simultaneously regulating the known BRCA1 activators ERα, IGF1R, SP1 and the repressor ID4, including a feedback regulation of miR-335 expression by estrogens [80]. Its overexpression causes decreased cell viability and increased apoptosis, while other findings show it to negatively regulate the HGF/c-Met pathway, thus affecting cell scattering, migration, and invasion [81].
Another downregulated miR in BC is miR-126 [82]. Its targets include VEGFA and PIK3R2 [83]. It also reduces trastuzumab resistance by targeting PIK3R2 and regulating the AKT/mTOR signaling pathway [84] and controls cell invasion by targeting ADAM9 [85].
Tumor suppressor miR-199a/b-3p inhibits migration and invasion of BC cells by downregulating the PAK4/MEK/ERK signaling pathway [86]. Overexpression of miR-199a-3p targets the c-Met and mTOR pathways, increases doxorubicin sensitivity and causes G1 phase arrest, thus reducing cell invasion and promoting doxorubicin-induced apoptosis [87]. This miR also confers resistance to cisplatin treatment by downregulating TFAM [88] and, at the same time, promotes BC development and metastasis under hypoxic conditions by controlling the regulatory axis consisting of HIF-1, SNHG1, and TFAM [89]. The same study mentioned above [87] also shows that in TNBC patients, additional circulating miR (i.e., miR-19a/b-3p, miR-25-3p, miR-22-3p, miR-210-3p and miR-93-5p) are deregulated as well and control several molecular pathways involved in drug resistance, making them amenable to be used as BC biomarkers, together with let-7a-5p, miR-100-5p and miR-101-3p, identified in another study [90].
Tumor suppressor miR-101 is downregulated as well in BC, and its targets include POMP, Stmn1, DNMT3A, EYA1, VHL, SOX2, Jak2 and MCL-1 (reviewed in [91]). For this reason, it plays a major role in the control of several cancer-related cellular processes, such as proliferation, apoptosis, angiogenesis, drug resistance, invasion, and metastasis. Overexpression of miR-101-3p can inhibit the migration of BC cells into the brain endothelium, a frequent and late event in BC patients, by inducing COX-2/MMP1 signaling, which can degrade the inter-endothelial junctions (claudin-5 and VE-cadherin) [92]. Jiang and collaborators showed that the suppression of the oncogene EZH2 in BC by miR-101-3p is potentiated in the presence of syn-cal14.1a, a synthetic peptide derived from Californiconus californicus (a sea snail), thus inhibiting cell migration, invasion, and proliferation [93]. Additional data come from the work of Toda and collaborators, who performed an RNA-sequence-based microRNA expression signature in BC and identified other dysregulated miRs in BC (e.g., miR-99a-5p/-3p, miR-101-5p/-3p, miR-126-5p/-3p, miR-143-5p/-3p, and miR-144-5p/-3p) and found that miR-101-5p controls the expression of seven putative oncogenes (i.e., HMGB3, ESRP1, GINS1, TPD52, SRPK1, VANGL1 and MAGOHB) [94].
Finally, miR-9 is known to exert critical functions in the initiation and progression of BC. Its upregulation—together with that of miR-221/222, miR-373 and miR-10b—is linked to highly malignant invasive EMT and cancer stem cell production [95]. Conversely, its downregulation can lead to improved overall survival, smaller tumors, earlier stages, and ER-positive cancers due to the enrichment of estrogen response genes [96]. Gwak and collaborators showed that miR-9 is highly expressed in HER2+ and TNBC subtypes compared with luminal subtypes, tumors with a high tumor stage or histologic grade, and tumors displaying the CD44+/CD24− phenotype, vimentin expression, and E-cadherin loss [97]. Interestingly, Shen and collaborators showed miR-9-5p, together with miR-195-5p and miR-203a-3p, to be a part of the extracellular vesicle (EV)-encapsulated miR (enabling cancer cell–cell communication in tumor pathogenesis and response to therapies) excreted upon docetaxel treatment [98]. As for its targets, genes identified so far include FOXO1 [99], STARD13 [100], LIFR [101], elf5A2 [102], HMGA2, EGR1, and IGFBP3 [68] and PDGFRbeta [103]. In turn, its expression is activated by MYC and MYCN [104,105].
A summary of the miR involved in BC and their function is summarized in Table 3.
Table 3. List of miR playing a direct role in BC. Target genes are those for which the miR/mRNA interaction is direct (usually, at the mRNA 3′ UTR), thus indirect interactions (e.g., other proteins of the same metabolic axis) are not reported in this table; additional miR studied only as BC biomarkers are collectively reported in the bottom row; see text for additional explanations. Abbreviations: n/a—data not available.
| miR Name | Target Gene(s) | Affected Cellular Functions | Refs |
|---|---|---|---|
| miR-21 | PTEN | drug resistance | [65,66] |
| miR-21 | LZTFL1 | proliferation and metastasis | [67] |
| miR-21 | IGFBP3 TPM1 PCD4 TGF-β1 |
proliferation, metastasis, epithelial-to-mesenchymal transition (EMT), apoptosis | [68] |
| miR-106a | RAF-1 | invasion and proliferation | [69] |
| miR-106a | P53 BAX RUNX3 Bcl-2 ABCG2 |
proliferation, colony-forming capacity, migration, invasion, apoptosis, sensitivity to cisplatin | [70,71] |
| miR-155 | TRF1 | telomere fragility | [72] |
| miR-141 | ANP32E | migration and invasion | [74] |
| let-7 | ERCC6 | proliferation, apoptosis | [78] |
| miR-335 | ERα IGF1R SP1 ID4 |
proliferation, apoptosis | [80] |
| miR-335 | c-Met | cell scattering, migration, and invasion | [81] |
| miR-126 | VEGFA PIK3R2 |
angiogenesis, tumor genesis and growth | [83] |
| miR-126 | PIK3R2 | trastuzumab resistance | [84] |
| miR-199a/b-3p | PAK4 | migration and invasion | [86] |
| miR-199a-3p | mTOR c-Met |
cell cycle progression, doxorubicin sensitivity, apoptosis | [87] |
| miR-199a-3p | TFAM | resistance to cisplatin | [88] |
| miR-199a-3p | TFAM | angiogenesis and metastasis under hypoxia | [89] |
| miR-101 | POMP Stmn1 DNMT3A EYA1 VHL SOX2 Jak2 MCL-1 |
proliferation, apoptosis, angiogenesis, drug resistance, invasion, metastasis | [91] |
| miR-101-3p | COX-2 | migration, metastasis | [92] |
| miR-101-3p | EZH2 | migration, invasion, proliferation | [93] |
| miR-101-5p | GINS1 | DNA replication | [94] |
| miR-9 | FOXO1 | proliferation, migration, invasion | [99] |
| miR-9 | STARD13 | EMT, metastasis | [100] |
| miR-9 | LIFR | metastasis | [101] |
| miR-9 | elf5A2 | resistance to doxorubicin | [102] |
| miR-9 | HMGA2 EGR1 IGFBP3 |
proliferation, metastasis, EMT, apoptosis | [68] |
| miR-9 | PDGFRβ | vasculogenesis | [103] |
| miR-200 | PDGFRβ | vasculogenesis | [103] |
| let-7a-5p miR-9-5p miR-10b miR-21 miR-22-3p miR-23b-3p miR-25-3p miR-29 miR-34a miR-93-5p miR-99a-5p/-3p miR-100-5p miR-101-3p miR-101-5p miR-126-5p/-3p miR-141-3p miR-143-5p/-3p miR-144-5p/-3p miR-145 miR-155 mir-181b1-5p miR-195-5p miR-199a-5p miR-200a miR-203 miR-203a-3p miR-205 miR-210-3p miR-221/222 miR-373 |
n/a | biomarkers | [73,76,87,90,94,95,97,98] |
All together, these data point to the involvement of several miRs in BC formation, development, metastasis, and drug resistance, showing that at the molecular level it is crucial to identify which pathways are altered and why, for example, the same gene may be deregulated because of the alteration of diverse miR. Knowing which miR is altered may greatly affect therapeutic approaches, especially in terms of avoiding cross-effects due to off-target actions. Thus, there is a necessity to identify (hopefully, all) the players in BC pathogenesis in a patient-specific way. In this perspective, the miR-125 family of miR has gained increasing relevance and attention in BC research, thanks to the numerous publications released over the last few years. In light of the numerous targets of these miRs and the multitude of pathways potentially altered inside the cell upon their dysregulation, in the next few years, miR-125 is likely to become central to understanding BC biology.
3. The miR-125 Family: Molecular Organization and Roles in Human Pathology
miR-125 is a highly conserved family of microRNAs whose members have also been found in nematodes (named lin-4 in 1993, the first miR described ever) [106]. The miR-125 family in H. sapiens includes three members, namely miR-125a, miR-125b-1 and miR-125b-2. The MIR125A gene maps to chromosome 19q13.41 [107], and miR-125a is part of a transcribed cluster of miR, together with miR-99b and let-7e [108]. The MIR125B1 gene maps to chromosome 11q24.1, and in this locus, it is part of a cluster including the LET7A2 and MIR100 genes [108,109]. These miRs are inside the third intron of the MIR100HG gene [110]. Finally, the MIR125B2 gene maps to chromosome 21q21.1, where it is included in a cluster together with the MIR99A and LET7C genes [108,109], inside the sixth intron of the MIR99AHG gene [110]. miR-125a and miR-125b differ only by a central diuridine insertion and a U-to-C change in miR-125a [111]. All members of the family show both 5p and 3p forms (Figure 1).
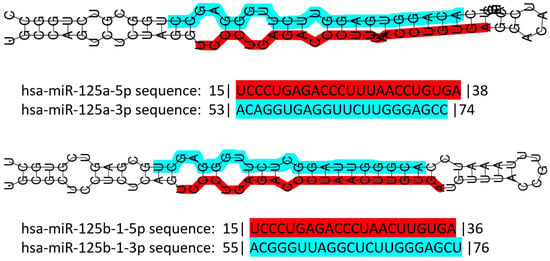
Figure 1. Schematic representation of human miR-125 illustrating the structure of the double-stranded pre-miR of both miR-125a and miR-125b, and their sequence. Color codes: red highlight for the 5p form, blue highlight for the 3p form. Numbers before and after the sequences indicate the number of nucleotides trimmed away from the mature miR. Data retrieved and partially modified from miRTarBase v9 update 2022 [112,113].
The miR-125 family is involved in several cell metabolic pathways controlling differentiation, proliferation, apoptosis, metastasis formation, drug resistance and immune system function because of the targeting of mRNAs related to these cellular processes [114] (Figure 2). miR-125 molecules have a complex behavior inside the cell, which mirrors their expression pattern in different tissues/cell types [112,113], their ample variety of targets [112,113], the intracellular role of their targets, and the way miR and mRNAs are either up- or down-regulated upon expression.
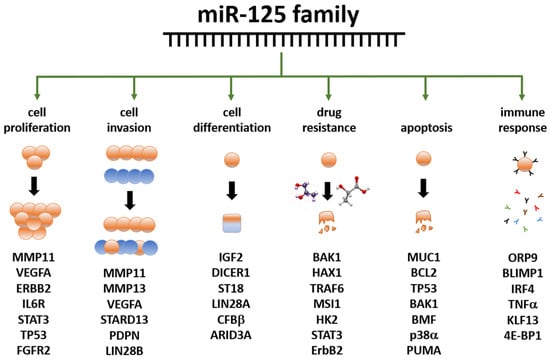
Figure 2. Summary of the main functions exerted by the miR-125 family members in human biology. Examples of miR-125 target genes are reported below each function. Data partly retrieved from [114,115] The listed genes at the bottom are involved in the control of the cellular functions below the green arrows; they are controlled, either directly or indirectly, by miR-125 family members, either by up- or down-regulation. Further details have been laid out in the article’s body.
The role of miR-125 family members has been extensively demonstrated in the muscle. It interacts with insulin-like growth factor II (IGF-II) to regulate myoblast differentiation in vitro and muscle regeneration in vivo [116], and with TRAF6 to prevent atrophy [117]. It is also involved in the proliferation and migration of vascular smooth muscle cells induced by platelet-derived growth factor BB [118]. In cardiac muscles, miR-125 participates in the development of the heart in embryonic mammals (reviewed in [119]); it regulates muscle-enriched transcription factors in cardiac and skeletal myocytes [120]; it can modulate cardiac progenitor cell proliferation and migration potential [121]; and it regulates cardiomyocytes proliferation and apoptosis under oxidative stress conditions [122]. Cardiac-specific miR-125b deficiency has recently been shown to induce perinatal death and cardiac hypertrophy [123].
miR-125 is one of the most abundant microRNAs in the central nervous system (CNS) in both mice and men [124]. In humans, miR-125b promotes neuronal differentiation in human cells by repressing at least ten target mRNAs involved in those pathways [125,126]. It also regulates dendritic spine morphology and synaptic maturation [127], it is implicated in synaptic plasticity [128], promotes astrogliogenesis, and is involved in astrogliosis and glial cell proliferation [129]. Its deregulation has also been linked to CNS tumor formation and growth, such as pediatric low-grade glioma [130]; it regulates cell growth arrest and apoptosis of human neuroblastoma- and medulloblastoma-derived cell lines [131,132]; it inhibits cell apoptosis through p53 and p38MAPK-independent pathways in glioblastoma cells [133]; and, in glioma, it targets BMF [134].
In the immune system, miR-125 regulates hematopoiesis, inflammation, and immune cell function. miR-125a controls stem cell homeostasis during hematopoiesis [135,136,137,138] and plays a role in immune cell identity [138]. miR-125-5p targeting IL-6R regulates macrophage inflammatory response and intestinal epithelial cell apoptosis in ulcerative colitis through the JAK1/STAT3 and NF-κB pathways [139]. miR-125b-1-3p is expressed in hMSCs-Ad exosomes and can promote T lymphocyte apoptosis and alleviate atherosclerosis (AS) by down-regulating BCL11B expression, thus providing potential molecular targets for the clinical treatment of AS [140].
All together, these data emphasize the multiple roles of miR-125 family members in cell proliferation and differentiation in numerous body locations.
4. miR-125 and Cancer
Studying the role of miR-125 in cancer is an important research area; beyond the above-mentioned tumors of the CNS, this noncoding RNA is indeed deregulated in several other tumors [141]. Additional organs affected by miR-125-related cancers include the ovary, bladder, liver, skin, bone, lung, pancreas, prostate, thyroid, stomach, colon and kidney. A summary of the cancers affecting human organs, known miR-125 targets and related bibliographic references are reported in Table 4.
Table 4. Summary of the affected organs and mRNA targets of miR-125 family members in human cancers, except for BC reported in the next section. Abbreviations: CNS—central nervous system; refs—references; n/a—data not available in the cited reference(s).
| miR | Organ | Target(s) | Notes | Refs |
|---|---|---|---|---|
| 125 | CNS | n/a | deregulated, pediatric | [130] |
| 125 | CNS | n/a | deregulated | [131,132] |
| 125 | CNS | p53, p38MAPK | none | [133] |
| 125 | CNS | BMF | none | [134] |
| 125a | ovary | n/a | EMT negative regulator | [144] |
| 125b | ovary | BCL3 | none | [145] |
| 125b | ovary | n/a | serum biomarker | [146] |
| 125b | bladder | E2F3 | none | [147] |
| 125b | bladder | n/a | urine biomarker | [148] |
| 125-3p | bladder | n/a | hypoxia regulated | [149] |
| 125 | bladder | n/a | survival predictor | [150] |
| 125a | liver | MMP11, VEGF | none | [151] |
| 125b | liver | Mcl-1, IL6R | none | [152] |
| 125b | liver | Lin28B2 | none | [153] |
| 125 | liver | Pokemon | none | [154] |
| 125 | liver | TRAF6 | none | [155] |
| 125 | liver | hexokinase II | none | [156] |
| 125 | liver | FOXM1 | none | [157] |
| 125 | skin | NCAM | none | [158] |
| 125 | skin | c-Jun | none | [159] |
| 125b | skin | MMP13 | none | [160] |
| 125b | skin | STAT3 | none | [161] |
| 125 | skin | n/a | deregulated | [162] |
| 125b | bone | STAT3 | none | [163,164] |
| 125 | bone | ErbB2 | none | [165] |
| 125 | bone | BAP1 | none | [166] |
| 125 | lung | n/a | survival predictor | [167] |
| 125 | lung | EGFR | none | [168] |
| 125 | lung | HER2 | trastuzumab resistance | [169] |
| 125 | lung | MMP13 | none | [170] |
| 125 | pancreas | n/a | deregulated | [171,172] |
| 125 | pancreas | NEDD9 | none | [173] |
| 125 | prostate | n/a | deregulated | [174,175,176] |
| 125 | prostate | BAK1 | none | [177] |
| 125 | prostate | p53, PUMA | none | [178] |
| 125b | thyroid | Foxp3 | cisplatin sensitivity | [179] |
| 125b | stomach | PPP1CA-Rb | none | [180] |
| 125a-5p | colon | BCL2, BCL2L12, MCL1 | none | [181] |
| 125b | kidney | n/a | survival predictor | [182] |
5. Role of miR-125 in BC
A relatively small amount of research is currently available on the role of miR-125 in BC. The reports showing its altered expression in these malignancies started to be published more than 20 years ago, and the research is still running in search of an affordable diagnostic panel based on this noncoding RNA [183,184,185,186,187,188]. Among the targets first identified, it is worth mentioning ERBB2 and ERBB3 [189] mRNA. In 2011, Zhang and colleagues demonstrated the action of miR-125b on the regulation of the ETS1 proto-oncogene in human invasive BC [190]. Rajabi et al. found that miR-125b, downregulated in BC, can reduce the expression of MUC1 (an oncoprotein), whose silencing causes DNA damage-induced apoptosis in cancer cells [191]. Tang and collaborators studied the effects of miR-125 deregulation on metastasis formation, finding that miR-125b induces metastasis by targeting STARD13 mRNA in MCF-7 and MDA-MB-231 BC cells [192], in contrast with the tumor suppressive action described before. Using the same BC cell lines, Metheetrairut and collaborators showed that forced expression of miR-125b results in radiosensitivity, as seen by reduced clonogenic survival, enhanced apoptotic activity and enhanced senescence post-ionizing radiation treatment. Moreover, re-expression of c-JUN in MDA-MB-231 cells promoted radioresistance and abrogated miR-125-mediated radiosensitization, suggesting that overexpression of miR-125b causes sensibilization to γ-irradiation and indicating this miR as a possible target for adjuvant therapy [193]. In contrast, Wang et al. found an association between miR-125b expression and chemoresistance [194], again indicating an oncogenic role for this miR. In line with these last results, Zhou and collaborators found that miR-125b confers the resistance of BC cells to paclitaxel through suppression of pro-apoptotic Bcl-2 antagonist killer 1 (Bak1) expression [195]. He and collaborators studied the expression of miR-125a-5p/3p and miR-125b in 143 pairs of BC and normal adjacent tissues, finding that miR-125a-5p and miR-125b were significantly down-regulated in BC tissue samples and that the expression level of miR-125a-5p was significantly higher in younger patients (<35 years) than in older ones, and a gradual reduction in miR-125a-5p expression was observed in BC tissue samples correlated to increasing age [196]. Recently, a paper showed the oncosuppressor role of miR-125b via the inhibition of proliferation, migration, and invasion of BC cells through targeting MMP11 protein expression [197]. A summary of the data reported above is illustrated in Table 5.
Table 5. Role of miR-125 in BC formation and development. In columns 2 and 4, reg. stands for regulation; an arrow pointing upwards means upregulation, while an arrow pointing downwards means downregulation; each arrow describes the miR/target regulation reported to its left. Note that in BC models, miR-125 members may be either up- or down-regulated, indicating either an oncogenic or oncosuppressive role for this molecule, in that context. Possible interpretations of these contradictory data are reported in the Discussion. In columns 3–4, n/a stands for ‘data not available’ or ‘not applicable’. In column 6, ‘blood samples’ means that circulating miR have been studied. In column 7, ref. stands for reference(s).
| miR | Reg. | Target | Reg. | Cellular Function | Cell Line | Ref. |
|---|---|---|---|---|---|---|
| miR-125a miR-125b |
↑ ↑ |
ERBB2 ERBB3 |
↓ ↓ |
migration invasion |
SKBR3 | [189] |
| miR-125b | ↓ | ETS1 | ↑ | proliferation | BC samples | [190] |
| miR-125b | ↓ | MUC1 | ↑ | apoptosis | BT-549 ZR-75-1 |
[191] |
| miR-125b | ↓ | STARD13 | ↑ | metastasis | MCF-7 MDA-MB-231 |
[192] |
| miR-125 | ↓ | n/a | n/a | radioresistance | MCF-7 MDA-MB-231 |
[193] |
| miR-125b | ↑ | n/a | n/a | chemoresistance proliferation apoptosis |
blood samples | [194] |
| miR-125b | ↑ | BAK1 | ↓ | chemoresistance apoptosis |
MDA-435 MDA-436 MDA-231 MCF7 SKBR3 |
[195] |
| miR-125a-5p miR-125b |
↓ ↓ |
n/a | n/a | age-dependent BC formation | BC samples | [196] |
| miR-125b | ↓ | MMP11 | ↑ | proliferation migration invasion |
T47D SKBR3 |
[197] |
6. Further Mining miR-125 Function in BC: Competing Endogenous RNA Networks (ceRNET)
A fundamental way to control gene expression through miRs has been elucidated in recent years, consisting of the so-called ceRNET. In fact, miRs have been shown to act as controllers of target mRNAs by altering their half-life or translation. However, they are also controlled, in many cases, by other long non-coding RNAs (lncRNA) or even other mRNA, which “sponge” miR through sequence homology, avoiding their interaction with mRNA targets [198]. In other words, lncRNA and mRNA compete for binding miR; these two molecules form a competing endogenous RNA (ceRNA) couple. If the lncRNA efficiently sponges the miR, then miR inhibitory action is not accomplished, and the target mRNA is regularly translated. In this case, the lncRNA, inhibitor of an inhibitor, has a function resembling that of an enhancer of gene expression. Hence, if the mRNA encodes an oncoprotein, the lncRNA has an oncogenic effect, while the miR has an oncosuppressive role. The same, with opposite effects, occurs in the case of the mRNA coding an oncosuppressor. The three molecules, taken together, form what is currently known as a regulatory axis, and the sum of many axes creates the ceRNET. Here, lncRNA and mRNA constitute the nodes of the network, while miR represent their connections. A growing number of research works have been published in recent years outlining the increasing structure and complexity of the ceRNET in BC (see [199] and references therein), including the action of pseudogenes in this phenomenon. In fact, Welch and collaborators found that 309 pseudogenes exhibit significant differential expression among BC subtypes, and their expression pattern allows recognizing tumor samples from normal samples and discriminating the basal subtype from the luminal and Her2 subtypes; of them, 177 transcribed pseudogenes possess binding sites for co-expressed miRs that are also predicted to target their parent genes [200]. Recently, in a work by Zhu and collaborators, the authors took advantage of the data available in the exoRbase database and derived it from the exosomes of human BC samples [201]. Their study allowed for the identification of a ceRNA network including 19 mRNA nodes, 2 lncRNA nodes, 8 circular RNA nodes, and 41 miR connections. KEGG enrichment analysis showed that differentially expressed mRNA in the regulatory network is mainly enriched in the p53 signaling pathway.
Research about portions of a miR-125-centered ceRNET has been expanding steadily. The miR-125 interactions with the mRNA described in the previous section are therefore likely to become axes of the growing BC ceRNET as well, as soon as the appropriate lncRNA is identified in the pathway. However, some axes have already been described, and some of them, being interconnected, can be used to build a basic version of this network (Figure 3).
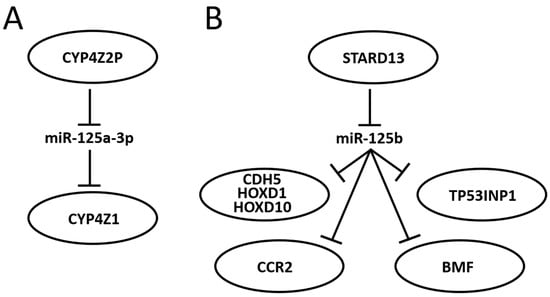
Figure 3. Schematic representation of two miR-125-centered ceRNET in BC. (A): a simple ceRNET having only one axis, where a pseudogene (CYP4Z2P) mRNA inhibits miR-125a-3p action on the target (CYP4Z1) mRNA by sponging it, thus enhancing CYP4Z1 expression. (B): a more complex ceRNET in which the interaction between miR-125b and STARD13 mRNA controls the expression of multiple target mRNA. See text for references and further explanations.
In 2004, Rieger and collaborators discovered a new human cytochrome P450 (CYP), termed CYP4Z1, which is specifically expressed in mammary gland and breast carcinoma [202]. They also found a transcribed pseudogene, named CYP4Z2P, that codes for a truncated CYP protein (340 amino acids vs. 505) with 96% identity to CYP4Z1. Both CYPs are highly expressed in BC, although the expression level of CYP4Z2P is approximately 20 times lower than that of CYP4Z1 in mammary tissues and barely expressed elsewhere. Later, it was shown that increased expression of CYP4Z1 promotes tumor angiogenesis and growth in human BC [203] and that CYP4Z2P 3′-UTR is involved in promoting BC angiogenesis through the VEGF/VEGFR2 pathway [204]. In 2015, Zheng et al. showed that the action of CYP4Z2P 3′-UTR is sponging several miRs, including miR-125a-3p, and that this pseudogene acts as a ceRNA with respect to CYP4Z1 mRNA, enhancing its expression levels [205]. They also showed that tumor angiogenesis is promoted by overexpression of the CYP4Z2P and CYP4Z1-3′UTRs, which significantly increased the activation of the ERK1/2 and PI3K/Akt pathways through the induction of their phosphorylation. The same group also showed a number of interactions later: (i) deregulation of these ceRNA also confers tamoxifen resistance in BC through the enhancement of the transcriptional activity of ERα via its phosphorylation dependent on cyclin-dependent kinase 3 (CDK3) [206]; (ii) downregulation of CYP4Z1 or CYP4Z2P through 3′-UTR binding promotes cell apoptosis, mirroring the functions and modulating the expression of human telomerase reverse transcriptase (hTERT) [207]; (iii) transcriptional factor six2 activates these CYPs ceRNET by directly binding to their promoters, thus activating the downstream PI3K/Akt and ERK1/2 pathways and consequently being involved not only in chemoresistance but also regulating the stemness of BC cells [208].
STARD13 (StAR-related lipid transfer domain protein 13, also known as deleted in liver cancer 2 protein (DLC-2)) is a Rho GTPase-activating protein (Rho GAP) that selectively activates RhoA and CDC42 and suppresses cell growth by inhibiting actin stress fiber assembly in hepatocellular carcinoma (HCC) [209]; this protein is ubiquitously expressed in normal tissues and downregulated in HCC. In mice, STARD13 promotes angiogenesis through the actions of RhoA [210]. Its role is well established in BC as well, where it acts as a tumor suppressor gene [211], regulates cell motility and invasion [212], endothelial differentiation [103], metastasis formation [213,214], cell migration [215], and apoptosis [216]. It has also been shown that STARD13 exerts its function in BC through its participation in many ceRNETs, such as the one involving a positive TGF-β/miR-9 regulatory loop mediated by the STARD13/YAP axis [217], the one involving hsa-miR-21-3p [218], or even the more complex network that involves five different miRs and that controls YAP/TAZ nuclear accumulation and transcriptional activity via modulation of Hippo and Rho-GTPase/F-actin signaling pathways [219]. A direct link between miR-125 and STARD13 expression has been described, too. Li and coworkers showed that CDH5, HOXD1, and HOXD10 encode putative STARD13 ceRNA and display concordant patterns with STARD13 in different metastatic potential BC cell lines and tissues; in addition, they also show that the 3′ UTR of STARD13 mRNA can bind miR-125b (and also miR-9 and miR-10b), indicating that this mRNA may participate in multiple pathways simultaneously [220], thus confirming their previous study about this interaction [192] and showing that the transcripts of the tumor suppressor genes CDH5, HOXD1, and HOXD10 inhibit BC metastasis in vitro and in vivo by competing with STARD13 mRNA for these three miR. Interestingly, CDH5, HOXD1 and HOXD10, along with STARD13, are BC players also in a different ceRNET, competing for a different set of miRs, indicating that STARD13′s role in BC is very complex. In 2017, Hu et al. discovered another ceRNET axis in which STARD13 and miR-125b control CCR2 (cysteine–cysteine chemokine receptor 2) expression levels [213]. In this case, the authors found that the CCR2 3′ UTR harbors three miR-125 binding sites that both inhibit MDA-MB-231 and MCF-7 cell metastasis by repressing epithelial-mesenchymal transition (EMT) in vitro and suppress BC metastasis in vivo through competition with STARD13 in a miR-125b-dependent and protein-coding-independent manner. Another component of the same ceRNET is TP53INP1 (tumor protein p53-inducible nuclear protein 1). TP53INP1 is an antiproliferative and proapoptotic protein involved in cell stress response that acts as a dual regulator of transcription and autophagy and is modulated by p53 in response to stress; it also interacts with kinases HIPK2 and PKCδ, which phosphorylate p53, creating a positive feedback loop between p53 and TP53INP1 [221]. TP53INP1 is also involved in SPARC (secreted protein acidic and rich in cysteine)-mediated-promotive effects on cancer cell migration and metastasis [222]. In 2018, Zheng et al. found a ceRNA interaction between STARD13 and TP53INP1 mediated by competitively binding to miR-125b in BC [223]. In this case, STARD13 promotes upregulation of TP53INP1, causing the inhibition of BC cell metastasis through competitive binding to miR-125b thanks to the inhibition of SPARC gene expression. Later, Guo and co-workers also found a ceRNET axis in BC involving miR-125b, STARD13 and BMF (Bcl-2-modifying factor) mRNA [216]. BMF is a member of the BCL2 protein family and controls apoptosis in several cell types [224]. The authors [216] found that miR-125b directly binds the 3′ UTR and thus downregulates BMF expression, and that STARD13, sponging miR-125b, upregulates BMF in BC both in vitro and in vivo. All together, these results suggest novel therapies for BC treatment and aid in selecting adequate drugs, depending on the molecular biology of the tumor, from a perspective aiming at the goal of personalized medicine. Indeed, a recent study showed that tanshinone IIA (an effective component extracted from Salvia miltiorrhiza that regulates the stemness of tumor cells) attenuates this phenotype in BC cells by downregulating miR-125b levels and upregulating its target gene STARD13 expression, while miR-125b overexpression or STARD13 knockdown impairs the inhibitory effects of tanshinone IIA on the stemness of BC cells [225].
This entry is adapted from the peer-reviewed paper 10.3390/ncrna10020016
This entry is offline, you can click here to edit this entry!