A large body of research shows an association between higher body weight and low vitamin D status, as assessed using serum 25-hydroxyvitamin D concentrations. Vitamin D can be metabolised in adipose tissue and has been reported to influence gene expression and modulate inflammation and adipose tissue metabolism in vitro. However, the exact metabolism of vitamin D in adipose tissue is currently unknown. White adipose tissue expresses the vitamin D receptor and hydroxylase enzymes, substantially involved in vitamin D metabolism and efficacy. The distribution and concentrations of the generated vitamin D compounds in adipose tissue, however, are largely unknown. Closing this knowledge gap could help to understand whether the different vitamin D compounds have specific health effects in the setting of adiposity.
1. Introduction
Vitamin D is a fat-soluble vitamin that has been reported to have many health effects. The common and most well known are the beneficial effects of vitamin D on bone density and skeletal health [
1,
2]. Moreover, vitamin D supplementation has been reported to decrease mortality in cancer patients and ameliorate type 2 diabetes in vitamin D-deficient individuals [
3]. There are several risk factors for vitamin D deficiency, such as age [
4], decreased sun exposure and skin pigmentation [
5], the use of certain medications and increased adiposity [
6]. Recently, vitamin D has come under scrutiny in relation to COVID-19, with studies showing a relation between low vitamin D status and higher susceptibility to infection [
7] and a possible beneficial effect of vitamin D supplementation in individuals with low vitamin D levels [
8].
Although vitamin D has been the subject of research for decades, the full picture of vitamin D’s action in the setting of adiposity is still unclear. An unexplored area is the focus on the levels of other vitamin D compounds, in addition to the current status marker 25-hydroxyvitamin D (25(OH)D). Several of these vitamin D compounds have already been under scrutiny [
19,
20,
21], but many questions remain unanswered. The aim here is to provide a broad overview of the current evidence surrounding the different effects of vitamin D compounds, with a special focus on people with higher body weight. Even though not all people with higher body weight are metabolically unhealthy [
22], a growing part of the global population is diagnosed with metabolic syndrome [
23]. Interestingly, a recent study has shown the relevance of focusing on metabolic health status instead of body weight only.
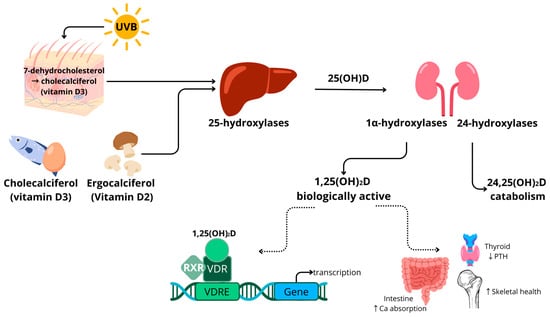
1.1. Vitamin D Metabolism
The precursors of 25(OH)D include vitamin D
3 (cholecalciferol), which is found in animal products, and vitamin D
2 (ergocalciferol), which is mainly present in mushrooms [
31,
32]. In humans, vitamin D
3 can also be generated in the skin by photosynthesis after UVB exposure of the precursor 7-dehydrocholesterol. Vitamin D
2 and vitamin D
3 follow the same metabolic pathway. The term vitamin D is used when vitamin D
2 or D
3 could be used interchangeably. The conversion of vitamin D to its different compounds is achieved through the action of cytochrome P450 enzymes, for example, vitamin D 25-hydroxylases (CYP2R1, CYP27A1 and CYP2J2), 1α-hydroxylase (CYP27B1) and the aforementioned CYP24A1. The activity of these enzymes is partially dependent on magnesium as a cofactor [
33,
34]. Hydroxylase enzymes are mainly expressed in the liver and kidney, as well as in other tissues and cell types, such as testes, blood cells, immune cells and adipose tissue [
30]. Vitamin D is converted in the liver (25 hydroxylases) and kidneys (1α hydroxylases) into bioactive 1,25-dihydroxyvitamin D (1,25(OH)
2D). Vitamin D is catabolised by 1,25-dihydroxyvitamin D hydroxylase (CYP24A1) to 24,25-dihydroxyvitamin D (24,25(OH)
2D) or 1,24,25-trihydroxyvitamin D (1,24,25(OH)
3D) [
30]. In circulation, vitamin D compounds are bound to the vitamin D binding protein (DBP) and albumin.
A schematic overview of vitamin D metabolism is provided in
Figure 1. Vitamin D binds to the vitamin D receptor (VDR), which is expressed in almost all human tissues, including adipose tissue [
35]. After binding, the VDR forms a complex with the retinoid X receptor (RXR). This complex is transferred to the nucleus, where it acts as a transcription factor. Many genes have a vitamin D response element (VDRE) in their promotor regions. Target genes are related to inflammatory processes and cell differentiation, in addition to bone and calcium metabolism [
36]. The transcription regulation by vitamin D is reported to be highly tissue-specific.
Figure 1. Schematic overview of vitamin D metabolism in humans. ↑ indicates an increase, ↓ indicates a decrease. Abbreviations: ultraviolet light short wavelength (UVB); calcidiol (25(OH)D); calcitriol (1,25(OH)2D); retinoid X receptor (RXR); vitamin D receptor (VDR); vitamin D response element (VDRE); parathyroid hormone (PTH); calcium (Ca).
Vitamin D metabolism is tightly regulated by parathyroid hormone (PTH) concentrations and is linked to calcium levels [
37]. Hypocalcaemia triggers the release of PTH, which, in turn, stimulates the activation of 25(OH)D to 1,25(OH)
2D in the kidneys by CYP27B1. The compound 1,25(OH)
2D increases intestinal calcium absorption, contributing to the normalisation of serum calcium levels and skeletal health, which is a major common health effect of vitamin D [
1].
1.2. Vitamin D in Adiposity
Two meta-analyses demonstrated a significantly higher prevalence of low vitamin D levels (serum 25(OH)D < 30 ng/mL) in adults and children with higher body weight [
39,
40]. Currently, the major hypotheses for this association are the sequestration of fat-soluble vitamin D in an increased amount of adipose tissue in people with elevated body fat mass [
41] and the volumetric distribution of vitamin D over a larger amount of body mass [
42]. Logically, this leads to the hypothesis that weight loss could contribute to achieving vitamin D sufficiency in people with higher body weight. However, the findings are currently inconclusive. Blum et al. showed a correlation between serum vitamin D
3 and WAT vitamin D
3 levels [
43]. They measured a mean vitamin D
3 content in WAT of 102.8 ± 42 nmol/kg in people with a mean body mass index (BMI) of 50.6 kg/m
2. This means that serum vitamin D could significantly improve upon weight loss if all vitamin D is released from adipose tissue. However, Himbert et al. reviewed the effect of surgery and diet-induced weight loss on serum 25(OH)D concentrations, and they concluded that weight loss did increase serum 25(OH)D levels but that this is often not enough to reach sufficient vitamin D concentrations [
44].
The expression and protein secretion of the hydroxylase enzymes involved in the conversion of vitamin D have been reported in many tissues, including WAT [
47]. Human adipocytes were shown to express CYP27B1, as well as CYP24A1 [
48]. The expression of the enzymes CYP2R1 and CYP2J2 was also shown in human adipose tissue [
49]. Jonas et al. demonstrated an increased CYP27B1 expression in WAT of people with higher body weight (BMI > 40 kg/m
2) [
50]. In another study, however, the expression of CYP2J2 and CYP27B1 was decreased in women with higher body weight (BMI > 35 kg/m
2) [
49]. These contradicting results illustrate that more research is necessary to understand the link between vitamin D metabolism and adipose tissue. However, hypotheses do exist; for example, the expression of CYP24A1 in adipose tissue could lead to an increased degradation of 1,25(OH)
2D in people with higher body weight. This effect could be aggravated by the concomitant downregulation of CYP2R1, another 25-hydroxylase, which was seen in a mouse model of obesity [
51]. Such changes in hydroxylase enzyme expression could be a partial explanation for the decreased vitamin D status in people with higher body weight and underline the link between adipose tissue and vitamin D metabolism.
In addition to hydroxylase enzymes, adipocytes express the VDR [
48,
49], meaning that vitamin D compounds binding the receptor can have a direct local effect on gene transcription in adipose tissue. Adipose VDR is reported to be increased in obesity [
50]. A genomic study showed that the rs3782905 single-nucleotide polymorphism (SNP) in the VDR gene is associated with differences in adiposity [
52], which indicates a relation between vitamin D signalling and the development of adipose tissue. However, the impact of different VDR genotypes on the development of adiposity is not supported by all data [
53]. Together, these findings on hydroxylase enzymes and VDR expression indicate once again that vitamin D metabolism is linked to adipose tissue mass. The relations between WAT and vitamin D metabolism in people with higher body weight are depicted in
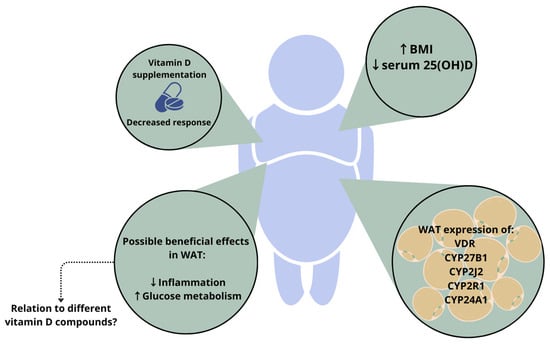
Figure 2.
Figure 2. Depiction of the relation between vitamin D status and white adipose tissue (WAT) mass in people with higher body weight. The response to vitamin D supplementation is altered in people with higher body weight, and an increased BMI is associated with lower serum 25(OH)D levels. In addition, white adipocytes express the VDR and hydroxylase enzymes. Vitamin D can have a beneficial effect on processes in WAT; however, it is unknown whether these effects are mediated by specific vitamin D compounds. ↑ indicates an increase, ↓ indicates a decrease. Abbreviations: BMI, body mass index; VDR, vitamin D receptor; WAT, white adipose tissue; 25(OH)D, 25-hydroxyvitamin D.
Body weight has also been reported to influence the response to vitamin D supplementation [
54]. A recent meta-analysis reported that vitamin D
3 supplementation is more effective than vitamin D
2 at raising serum 25(OH)D concentrations. However, the authors reported that baseline status needs to be considered and that this seems to be less relevant for people with higher body weight, since having a BMI > 25 k/m
2 reduced the difference in response to vitamin D
3 and vitamin D
2 supplementation [
55].
In this context, it is also important to consider the differences between the different types of adipose tissue. Visceral adipose tissue (VAT) expansion seems to have a more unfavourable effect on metabolic health than an increase in subcutaneous adipose tissue (SAT) [
60,
61].
Of note, the rapid expansion of WAT in people with higher body weight can lead to an inflammatory response in adipocytes and adipose tissue macrophages [
17]. These changes in adipose tissue function induce an increase in insulin resistance, which is a marker of a metabolically unhealthy phenotype in people with higher body weight [
23,
65]. Considering the reported association of vitamin D with inflammation [
66,
67] and glucose metabolism [
15,
16], differences in vitamin D availability or action in adipose tissue could play a crucial role in this context.
2. Relevance of Different Vitamin D Compounds
2.1. Free Vitamin D Compound
The majority of vitamin D is bound to the vitamin D binding protein (DBP). Megalin/cubulin receptors enable the entry of the vitamin D-DBP complex into cells [
68]. The proportion of free vitamin D is a very small fraction in comparison to that of protein-bound vitamin D, but it can move freely over cell membranes [
69]. According to the free hormone hypothesis, this makes free vitamin D the most biologically active vitamin D compound [
69,
70,
71], and, thus, this would make it very relevant in relation to the health effects of vitamin D.
Schwartz et al. performed a large cross-sectional study, in which they measured total serum 25(OH)D and free 25(OH)D levels in people with liver cirrhosis, pregnant women and healthy controls [
72]. They observed that the levels of free 25(OH)D were significantly higher in people with liver disease, most likely due to a decreased capacity to produce DBP.
A bariatric surgery study showed that weight loss led to differential changes in the serum levels of free 25(OH)D and total 25(OH)D [
74]. The serum levels of total 25(OH)D did not significantly increase when assessed one-year post-surgery, whereas a rise in the serum levels of free 25(OH)D could be observed. This could indicate that free vitamin D is the main storage compound of vitamin D in adipose tissue. However, it should be noted that there are several other mechanisms that could explain this differential effect, such as changes in serum protein levels or the expression of the vitamin D binding protein after weight loss. In a study comparing lean women to women with higher body weight (BMI = 39.1 ± 4.6 kg/m
2), DBP levels were higher and free vitamin D levels were lower in the latter group [
75]. The increased DBP levels could potentially have an effect on the availability of free vitamin D in tissues. Of note, the free vitamin D levels in this study were calculated rather than quantified with immunoassays or mass spectrometry.
2.2. 1,25-Dihydroxyvitamin D (1,25(OH)2D)
The 1,25(OH)
2D compound is generally considered the active form due to its high affinity for the VDR [
77].
With regard to the anti-inflammatory effects of 1,25(OH)
2D, NFkB signalling has been implicated as a potential mechanism, since vitamin D has been reported to decrease NFkB expression [
12] and to block NFkB translocation to the nucleus [
13,
78,
79], thus decreasing the transcription of pro-inflammatory cytokines. Even though these findings directly link 1,25(OH)
2D to decreased inflammation in vitro, there are contradicting reports of the effects of vitamin D supplementation on inflammatory markers in vivo. Wamberg et al. [
80] observed a diminishing effect on inflammation in an adipocyte cell line treated with 1,25(OH)
2D
3. However, this effect was not reproduced in primary human adipocytes, and it was not reflected in circulating inflammatory markers in vivo [
80].
Adipokines are messenger molecules produced by adipocytes. They affect energy metabolism, insulin resistance, inflammation and blood pressure and are strongly related to metabolic health [
84,
85]. Two well-known examples are leptin and adiponectin. Leptin is known as the satiety hormone, which, in healthy conditions, supresses food intake, regulates energy metabolism and acts as a negative feedback loop upon adipose tissue accumulation [
86,
87]. However, in people with higher body weight, leptin resistance frequently occurs [
88,
89,
90]. Adiponectin is an adipose-tissue-derived hormone with health-promoting effects. It has been reported to increase insulin sensitivity and have anti-inflammatory properties [
91]. Adiponectin levels are often decreased in people with increased body fat [
92].
The effects of 1,25(OH)
2D exposure on adiponectin have also been reported, e.g., decreased adiponectin production in human primary adipocytes [
99]. Accordingly, a negative association between adiponectin levels and serum 25(OH)D was observed in humans [
97]. However, this effect was only reported in a subgroup of participants with a BMI > 25 kg/m
2. In another study, no association between serum 25(OH)D and adiponectin was reported [
100]. And yet another study found a positive correlation between 25(OH)D and adiponectin levels [
96]. Two trials did not show an effect of vitamin D on adipokine levels [
101,
102].
2.3. 24,25-Dihydroxyvitamin D (24,25(OH)2D)
The 24,25(OH)
2D compound is a clearance product in the vitamin D metabolic cascade. The ratio of 24,25(OH)
2D to 25(OH)D has been suggested to be an alternative marker of vitamin D status [
103]. This ratio takes vitamin D metabolism and feedback loops into account and is not dependent on the level of DBP [
104,
105]. When assessing other vitamin D compounds, Binkley et al. described the predictive value of 24,25(OH)
2D for the response to vitamin D supplementation [
20]. In this study, the ratio of 24,25(OH)
2D/25(OH)D did not predict the response to vitamin D supplementation, contradicting earlier suggestions of 24,25(OH)
2D/25(OH)D being an improved status marker [
103,
106]. However, the sample size was relatively small (
n = 62) and consisted of postmenopausal women only. These data indicate that the measurement of 24,25(OH)
2D could have an additional clinical relevance to measuring only serum 25(OH)D [
106].
2.4. C-3 Epimers of Vitamin D Compounds
In vitamin D metabolism, a parallel C-3 epimerisation pathway introduces additional vitamin D compounds. The most important of these is the C-3α isomer of 25(OH)D, which results from the reversal of the stereochemical configuration of the –OH group at C-3 (3β
→3α). It is important to measure this compound for two reasons. Firstly, the 3α epimer has been reported to lead to the overestimation of vitamin D status with current detection methods, such as immunoassays and MS assays, if it is not properly separated [
109]. The extent of this additional contribution depends on physiological and pathological conditions and age; e.g., 3α levels are naturally higher in newborns, especially in those born prematurely [
110,
111].
With respect to body composition, a negative correlation was reported for serum 25(OH)D concentrations and fat mass in infants at 12 months in a vitamin D supplementation trial [
113]. The authors reported that serum 25(OH)D was a predictor of lean mass and fat mass in regression models. This correlation was only present at the age of three months for 3α-25(OH)D, even though 3β-25(OH)D was associated with all time points. In a Thai national health survey, the relative amount of 3α-25(OH)D
3 was associated with age, sex and living conditions [
114]. Males and people living in rural areas had higher relative concentrations of serum 3α-25(OH)D
3. The survey observed a negative correlation between BMI and 3β-25(OH)D, but not 3α-25(OH)D. Another study that included women with polycystic ovary syndrome (PCOS) and healthy controls did not observe differences between the two groups in 3β and 3α-25(OH)D
3 [
115].
This entry is adapted from the peer-reviewed paper 10.3390/nu16020231