Membrane transporters are proteins embedded in plasma membranes and organellar membranes [
1]. These proteins are distributed in various tissues or cells and can improve the efficiency of plants in utilizing water and mineral elements [
2,
3] and transporting sugars to provide energy for plants [
4,
5]. They are also involved in the absorption, transportation, and detoxification of heavy metal substances by plants [
6]. Recent studies have shown that complex interactions occur between many membrane transport proteins and ROS in plants. Membrane transporters can be activated by ROS signaling to perform related transport functions [
7]. In turn, the transport of ions, sugars, hormones, amino acids, and other substances by membrane transporters can trigger a series of physiological metabolic reactions in plants, which enhance antioxidant enzyme activity, scavenge excess ROS, and regulate plant tolerance under abiotic stress [
8,
9,
10,
11]. Under abiotic stress, ROS accumulate in different forms (
1O
2, O
2•−, H
2O
2, and
•OH) in the cytosol and in various plant organelles [
12,
13,
14]. Excessive ROS can interfere with cell homeostasis, disrupt lipids and DNA, and ultimately lead to cell apoptosis [
15,
16,
17].
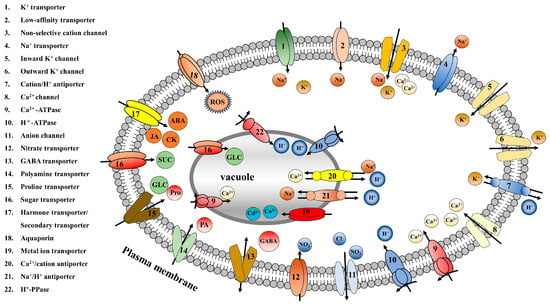
The study of membrane transporters can be traced back to the 1950s. Subsequently, membrane transporters were found to exist widely in plants and animals. Membrane transporters are embedded in the plasma membranes of cells and various organellar membranes and can be classified into different types based on their transport characteristics for different substances (
Figure 1). These different types of membrane transporters perform different functions. Ion transporters can transport a variety of ions, including Na
+, K
+, Ca
2+, H
+, and Cl
−, as well as heavy metal ions such as Ni
2+ and Cd
2+, regulating intracellular ion concentrations and maintaining the cellular pH balance. Sugar transporters can transport sucrose, fructose, glucose, and various sugar alcohols to provide energy for plants. Amino acid transporters, hormone transporters, and other secondary metabolite transporters are involved in the transport of related substances and regulate various metabolic reactions in plants, playing key roles in research on the application of exogenous substances. These membrane transporters exist in plants as carrier proteins and channel proteins. Through their absorption and transport functions, they increase the levels of beneficial nutrients within cells, playing important roles in improving plant growth and development and enhancing plant tolerance to abiotic stress [
19,
20,
21,
22].
Figure 1. Classification diagram of ion transporters, sugar transporters, amino acid transporters, hormone transporters, and secondary metabolite transporters in plants. Arrow pointing represents the direction of transportation.
2. Ion Transporters
2.1. Na+ Transporters
Na
+ is the most abundant type of cation in extracellular fluid, playing a role in maintaining cellular water and the acid–base balance [
23]. There are two main types of Na
+ transporters in plants. The first type is located on the plasma membrane and controls the transport of Na
+ across the plasma membrane. The influx of Na
+ is controlled by high-affinity K
+ transporters (HKTs) [
24,
25,
26,
27,
28,
29], low-affinity transporters (LCTs), nucleotide-gated channels (CNGCs), and ionotropic glucose receptor (GLR) channels [
30,
31,
32]. The efflux of Na
+ is controlled by salt overly sensitive 1 (SOS1) [
33]. The second type is located on the vacuolar membrane and controls the transport of Na
+ across the vacuolar membrane. Na
+/H
+ antiporters (NHXs) control the transport of Na
+ from the cytosol to vacuoles through the exchange of Na
+ and H
+ [
34], which reduces the Na
+ content in the cytosol and increases plant tolerance. In addition, some studies have indicated an interaction relationship between NHXs and SOSs, but the specific underlying mechanism still needs further exploration.
2.2. K+ Transporters
K
+ is the main cation in intracellular fluids and plays an important role in promoting plant growth and development, enhancing photosynthesis and material synthesis within plants, and improving sugar and energy metabolism [
8,
35]. Due to the difference in K
+ concentration between soil and plants, the transport of K
+ requires energy [
36,
37]. There are many K
+ transporters in plants, including HKT, KT/HAK/KUP, AKT, two-pore channels (TPCs), and cation/H
+ antiporters. These transporters are distributed on the plasma membrane and vacuolar membrane and can transport K
+ under different conditions. In 1994, HKT was identified as a high-affinity K
+ transporter protein that is an alkaline cation transporter linking cytosolic osmotic homeostasis with plant tolerance under salt stress and contributing significantly to Na
+ transport [
27,
38]. The KT/HAK/KUP transporter family belongs to the amino acid polyamine–organocation superfamily, among which the HAK transporter has more obvious characteristics [
39,
40]. It regulates the transport of K
+ in low K
+ concentration environments and is involved in the redistribution of K
+ to maintain Na
+/K
+ levels [
41,
42]. The AKT family includes AKTs and KATs, which are K
+-channel proteins [
43]. There are four types of K
+-channel proteins, namely inward-correcting (K
in) channels, weakly-correcting (K
weak) channels, silent (K
silent) channels, and outward-correcting (K
out) channels [
37,
44,
45,
46,
47]. TPCs are located on the plasma and vacuolar membranes, and their main function is to regulate the transport of cytosolic K
+ to maintain normal Na
+/K
+. In addition, two types of cation/H
+ antiporters, CHX and KEA, can also provide additional K
+ transport capacity in high-concentration K
+ environments, but their transport mechanism is unclear [
48,
49].
2.3. Ca2+ Transporters
Ca
2+ is an essential nutrient for plants. Ca
2+ homeostasis is highly important for maintaining the integrity of the cell membrane structure and for maintaining intracellular enzyme activity [
50]. Like Na
+, Ca
2+ membrane transporters are located on the plasma membrane and control the transport of Ca
2+ across the plasma membrane. The influx of Ca
2+ is controlled by mechanosensitive channels (OSCAs), CNGCs, GLRs, TPCs, etc. [
51,
52]. OSCA1 can play a role in osmotic stress [
53], and TPC channels can specifically mediate the influx of Ca
2+ [
54]. The efflux of Ca
2+ is energy-dependent and is mainly achieved through Ca
2+-ATPase. In
Arabidopsis, the autoenriched Ca
2+-ATPase (ACA) genes ACA2 and ACA4 have been shown to control the efflux of Ca
2+ [
55,
56,
57]. Another type of Ca
2+ membrane transporter is located on the vacuolar membrane and controls the efflux of Ca
2+ from the cytosol to the vacuole; this process is mainly achieved through Ca
2+/cation antiporters [
58]. Ca
2+/Na
+ exchange (NCL) can transport Ca
2+ to the vacuole through the exchange of Ca
2+ and Na
+ [
59], and Ca
2+/H
+ exchange (VCX, CAX) can transport Ca
2+ to the vacuole through the exchange of Ca
2+ and H
+ [
60,
61].
2.4. H+ Transporters
Hydrogen atoms lose electrons to form H
+, which can regulate the pH inside plants, promote plant growth and development, and improve nutritional quality [
19]. H
+-ATPase and H
+-PPase are involved mainly in the transport of H
+ in cells. H
+-ATPases are divided into plasma membrane H
+-ATPases (PMAs) and vacuolar membrane H
+-ATPases (VMAs). PMAs can generate a proton gradient, which drives SOS1 to transport Na
+ [
62]. VMAs and V-H
+-PPases are located on the vacuolar membrane and are responsible for transporting H
+ from the cytosol to the vacuole [
63,
64]. V-H
+-PPases have higher activity in young tissues, while VMAs have higher activity during plant growth and maturity [
65]. These two types of transporters generate H
+ gradients on the vacuolar membrane, driving NHXs to transport Na
+ [
66,
67]. H
+ transporters play a crucial role in maintaining ion homeostasis and improving plant tolerance under abiotic stress through compartmentation.
2.5. Anion Transporters
Inorganic anions in plants include chloride (Cl
−) and nitrate (NO
3−) ions, which are regulated both inside and outside the cell by two anion channel proteins: slow anion channels (SLAC/SLAH) and chloride channels (CLC) [
68]. SLACs can regulate the distribution of anions in the xylem in the extracellular space [
69,
70]. CLCs regulate the transport of anions through their intracellular compartmentalization effect [
71]. In addition, aluminum-activated malate transporters (ALMTs) are distributed on the plasma and vacuolar membranes and are involved in the transport of Cl
− [
72,
73,
74,
75]. NTRs are located on the plasma membrane and rely on the H
+ gradient provided by H
+-ATPase for NO
3− transport [
76].
2.6. Other Ion Transporters
Metal ions such as Fe
2+, Zn
2+, and Mg
2+ are regulated by various membrane transporters in plants [
77]. Some membrane transporters have specificity for a single type of ion, while others can transport multiple types. Mg
2+ transporters (MGTs) are distributed in the roots and leaves of plants and are responsible for Mg
2+ transport. The iron nicotianamine transporter yellow-stripe-like 2 (OsYSL2) is responsible for the transport of Fe
2+ in plants [
78]. Metal tolerance proteins (MTPs) control the transport of Zn
2+ and are associated with Zn
2+ sensitivity and tolerance [
79]. Vacuole iron transporters (VITs) control the transport of Fe
2+, Zn
2+, Mg
2+ [
80], etc. In addition to these elements essential for plant growth and development, studies have shown that there are many toxic heavy metal ions in the soil environment. Membrane transporters play a crucial role in heavy metal ion scavenging, detoxification, soil improvement, and enhancement of plant tolerance to heavy metal stress. Cation diffusion facility (CDF) transporters are a type of cation/H
+ antiporter that can transport heavy metal ions such as Cd
2+, Co
2+, and Ni
2+ through the exchange of cations and H
+ [
81]. Iron-regulated transporters (IRTs) control the transport of Cd
2+ and Ni
2+ in plants [
82]. Natural resistance-associated macrophage proteins (NRAMPs) are located on the vacuolar membrane and transport Cd
2+ to the vacuole for chelation [
83]. ATP binding cassette (ABC) transporters are the most ubiquitous in plants and are currently the largest family of membrane transporters [
84]. Multidrug-associated proteins (MRPs) are ABC transporters that are involved in the transport of Cd
2+ in plants, but their specific mechanism is unclear [
85].
3. Sugar Transporters
Sugars are important components of plant cells and occur in the form of sucrose, fructose, glucose, starch, and other substances in plant cells. They are responsible for energy supply and signal transduction in plants. Sugar transporters ensure the long-distance distribution of sugars in cells and tissues and are involved in signal transduction for the perception of abiotic stress and environmental adaptation [
86]. There are three main types of sugar transporters in plants: sugar transporters (SUTs), sugar will be exported transporters (SWEETs), and monosaccharide transporters (MSTs) [
4]. SUTs are located on the plasma membrane and are only found in plants. These proteins have been identified in rice and
Arabidopsis and are responsible for the long-distance transport of sucrose in plants [
87]. SWEETs are distributed on both the plasma membrane and the vacuolar membrane and have been identified in plants such as rice,
Arabidopsis,
Camellia sinensis, and
Dianthus spiculifolius. They can passively transport sucrose, glucose, and fructose along concentration gradients [
87,
88,
89,
90]. MSTs belong to the major facility superfamily, which consists of seven subgroups: early response to dehydration (ERD6), sugar transporter proteins (STPs), plastic glucose transporter (pGlcT), inositol transporters (INTs), vacuum glucose transporters (VGTs), tonoplast sugar transporters (TSTs), and polymer/monosaccharide transporters (PLTs). The different subfamilies of MSTs are distributed in different locations, controlling the transport of sucrose, maltose, glucose, sugar alcohols, and other sugars and regulating various physiological functions in plants, such as sugar distribution and signal perception [
91,
92,
93,
94,
95]. Multiple sugar transporters can control sugar transport, and further research is needed to determine which of these transporters plays a major role in sugar transport in plants.
4. Amino Acid Transporters
Amino acids are key nutrients required by plants and play an important role in promoting plant photosynthesis and material metabolism and in enhancing plant tolerance. The amino acid transporter (AAT) family can be divided into two categories: the amino acid polyamine choline transporter (APC) family and the amino acid/auxin permease (AAAP) family [
96]. The APC transporter superfamily includes cation amino acid transporters (CATs), polyamine H
+ cotransporters (PHSs), and amino acid/choline transporters (ACTs). CATs control the bidirectional transport of GABA, glutamate, and aspartate between the cytosol and vacuoles [
97]. PHSs mainly play a role in polyamine transport [
98]. ACTs control the bidirectional transport of GABA between the cytosol and mitochondria [
99,
100]. AAAPs include amino acid permanence transporters (AAPs), lysine/histidine transporters (LHTs), proline transporters (ProTs), aromatic and neutral amino acid transporters (ANTs), putative auxin transporters (AUXs), GABA transporters (GATs), etc. [
101,
102,
103,
104,
105]. The AAAP family plays an important role in the transport of GABA, lysine, histidine, proline, and many other amino acids. Although studies on amino acid transporters have been reported for many years, many of them have not been studied in depth, and fully understanding the regulation of amino acids by transporters in plants is still highly challenging.
5. Other Compound Transporters
Compounds such as plant hormones and secondary metabolites can regulate plant growth and development. Transporter families such as ABC transporters, multidrug and toxic compound extrusion (MATE) transporters, purine uptake permease (PUP) transporters, and nitrate–peptide (NRT) transporters are involved in the transport of these compounds [
106]. Each of these transporter families performs different transport functions. The G-type ABC transporter mediates the transportation of abscisic acid (ABA), controls physiological responses such as stomatal closure and leaf temperature changes in plants, and increases plant tolerance. B-type and C-type ABC transporters are involved in the transport of berberine, anthocyanins, and other flavonoids in plant tissues [
107,
108]. MATE transporters can transport alkaloids, including nicotine, anabasine, and scopolamine, to enhance the chemical defense of plants against microorganisms and pests [
109]. PUP transporters can transport cytokinins to regulate the differentiation of plant roots and shoots. NRT transporters have been shown to play a role in the transport of various substrates, such as peptides, IAA, and GA [
110,
111,
112,
113,
114]. At present, the transport mechanisms of many hormones and other compounds in plants are still unclear and require further research.