Oogenesis is a developmental process leading to the formation of an oocyte, a haploid gamete, which upon fertilisation and sperm entry allows the male and the female pronuclei to fuse and give rise to a zygote. In addition to forming a haploid gamete, oogenesis builds up a store of proteins, mRNAs, and organelles in the oocyte needed for the development of the future embryo. In several species, such as Drosophila, the polarity axes determinants of the future embryo must be asymmetrically distributed prior to fertilisation. In the Drosophila oocyte, the correct positioning of the nucleus is essential for establishing the dorsoventral polarity axis of the future embryo and allowing the meiotic spindles to be positioned in close vicinity to the unique sperm entry point into the oocyte.
1. Anatomy and Development of the Drosophila Egg Chamber
A typical ovary comprises approximately 16 ovarioles, each representing an independent egg assembly chain with a tubular organisation. Each ovariole encloses at its tip a structure, the germarium, which is associated with germline and somatic stem cells, whose progeny becomes organized into ovarian follicles or egg chambers. The follicles exit the germarium and continue to develop through a process divided into 14 stages on morphological grounds as they move posteriorly within the ovariole (
Figure 1 and
Figure 2) [
1]. Throughout
Drosophila oogenesis in adult females, unlike in mammals, germ line stem cells produce a constant supply of new oocytes [
2,
3].
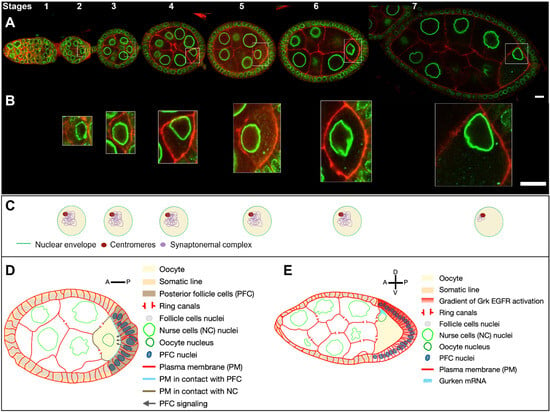
Figure 1. Drosophila egg chamber organisation and oocyte nucleus positioning from stage 1 to 7. (
A) Reconstitution of an ovariole from different ovarian follicles from stage 1 to stage 7. The nuclei are visualized by the expression of the
Drosophila importin-β i. e Female sterile (2) Ketel protein coupled to GFP (
Fs(2)Ket-GFP) (green) and cell membranes by the ubiquitous expression of the pleckstrin homology (PH) domain of the Phospholipase C protein coupled to RFP (
ubi-PH-RFP) (red) (scale bar 10 microns). (
B) Close-up of the nucleus in the oocyte, illustrating the evolution of nuclear positioning in the oocyte from stage 2 to stage 7 (scale bar 10 microns). (
C) Schematic representation of the internal nuclear organisation adapted from [
4]. At stages 2 to 4, the chromosomes reorganise to form a compact structure of condensed inactive chromatin called the karyosome. [
5,
6]. From stage 5 to 6, the euchromatic synaptonemal complex is disassembled. At stage 7, the synaptonemal complex persists at the centromeres. (
D) Schematic representation of a stage 6 ovarian follicle. The posterior follicles cells (dark brown) in contact with the plasma membrane at the posterior of the oocyte (blue dotted line) send a signal (black arrows) to the oocyte, which eventually leads to the migration of the nucleus (
B) and the reorganisation of the microtubule networks. (
E) Schematic representation of a stage 7 ovarian follicle. The nucleus has migrated to the intersection of the anterior and posterior plasma membranes of the oocyte. This position will define the dorsal pole of the oocyte. The nucleus is associated with
grk mRNA, which is translated and secreted locally. This creates an activation gradient for the EGF signalling pathway, which is responsible for establishing the dorsoventral polarity of the ovarian follicle [
7] and the future embryo [
8].
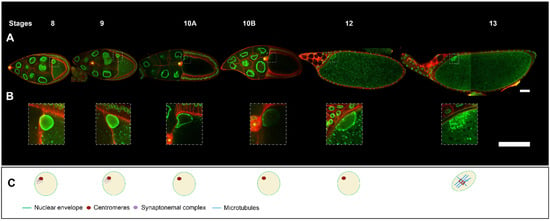
Figure 2. Drosophila egg chamber organisation and oocyte nucleus positioning from stage 8 to 13. (
A) Reconstitution of an ovariole from different ovarian follicles from stage 8 to stage 13. The nuclei are visualized by the expression of
Fs(2)Ket-GFP (green) and cell membranes by
ubi-PH-RFP expression (red) (scale bar 50 microns). (
B) Close-up of the oocyte nucleus, illustrating the evolution of nuclear positioning from stage 8 to stage 13. The border cells (yellow asterisk) which have delaminated from the anterior follicle cells at the end of stage 8, reached at stage 10 the anterior border of the oocyte and then migrated near the nucleus in the oocyte [
9] (scale bar 50 microns). (
C) Schematic representation of the intra-nuclear organisation adapted from [
4]. At stages 8 and 9, as at stage 7, the centromeric synaptonemal complex persists. At stage 10, chromosomes briefly decondense and transcription is upregulated [
5]. From stage 11 to 12, the chromosomes recondense. At stage 13, the germinal vesicle breaks down. Tubulin is recruited to the chromosomes and microtubules (MTs) begin to organise into a bipolar spindle. The oocyte maintains the metaphase 1 arrest configuration until it passes through the oviduct, triggering the end of meiosis I and the start of meiosis II.
Following an asymmetric division, a germline stem cell (GSC) produces another stem cell and a cystoblast, which undergoes four incomplete mitotic divisions to generate a cyst of 16 cells connected by cytoplasmic bridges or “ring canals” [
10]. Initially, two cells within the cyst with four ring canals initiate premeiotic development with the pairing of homologous chromosomes and the assembly of the synaptonemal complex along the chromosome arms [
11]. Then, at stage 1 in the germarium, one of these two cells differentiates into an oocyte, remaining in meiosis, while the 15 other cells of the cyst exit meiosis and eventually endoreplicate their genome to become nurse cells. The oocyte nucleus arrested in prophase of meiosis I, is transcriptionally quiescent or at least poorly active and the chromosomes are condensed in a structure named karyosome. Nurse cells become highly polyploids and provide the oocyte with all types of RNAs, proteins, and cellular organelles [
10]. Similarly to mammals, there is a long pause in meiotic prophase, between the pachytene stage where recombination occurs and the metaphase I stage where a meiotic spindle assembles at stage 13 [
4].
Nurse cells supply to the oocyte is closely linked to the microtubule (MT) cytoskeleton, through a polarized transport from the nurse cells to the oocyte [
12,
13,
14,
15]. MT organisation in the oocyte depends on several sources associated or not with the centrosomes [
16,
17,
18]. During the four incomplete divisions that ensure cyst formation, the 16 cells are linked to each other through the ring canals by a cytoplasmic structure known as the fusome, which combines cytoskeleton and vesicles [
10]. At the level of the fusome, the spectraplakin Shot, recruits the MT minus end stabilizer Patronin. During cyst formation, asymmetric fusome segregation results in one of the two pro-oocyte cells with four ring canals having more fusome material than the other. Together with the dynein MT motor, Shot and Patronin through an amplification process, create an MT enrichment in the pro-oocyte where the fusome is more abundant, which is required for oocyte specification [
19]. Therefore, the selection of the oocyte relies on the formation in the future oocyte of a noncentrosomal microtubule organizing center (ncMTOC) that organises a polarized MT network directing the dynein-dependent transport of cell fate determinants and centrosomes into the pro-oocyte [
19,
20,
21].
Centrosomes have a peculiar organisation in the cyst and in the developing oocyte. When the oocyte is determined, the centrosomes migrate from the 15 nurse cells through ncMTs towards the oocyte [
19,
21]. Because the oocyte is arrested in prophase of meiosis 1, the centrosomes are partially duplicated and between 15 and 32 centrosomes can be observed in the oocyte [
18]. During stages 5 to 6, the centrosomes cluster and migrate between the nucleus and the posterior pole of the oocyte where they are active and eventually migrate with the nucleus to an antero-dorsal position [
18,
22,
23]. The meiotic spindle is devoid of centrosomes. This is important because the centrosome of the future zygote is brought by the spermatozoid and the maternal centrosomes, if being maintained, would hinder the formation of the zygote. Hence, in the developing oocyte the centrosomes have disappeared by the end of oogenesis. Elegant experiments have shown the pericentriolar material is first gradually eliminated from the centrosomes in a stepwise manner before the centrioles disappear [
24].
An important issue is the asymmetric distribution during oogenesis of the determinants of the antero-posterior and dorsoventral polarity axes of the future embryo, which are essential for its segmentation and the formation of the different germ layers. This process relies on the asymmetric localisation of several mRNAs in the oocyte, with those of the
bicoid,
oskar (
osk), and
gurken (
grk) genes of particular importance [
25]. These mRNAs are transcribed in the nurse cells and transported in a MT-dependent manner into the oocyte through the ring canals (
Figure 1D) [
26]. Importantly, they are translated only when their transport is completed, ensuring the localisation of the encoded proteins is restricted to the area where the mRNAs are transported and anchored. The
bicoid mRNA is transported to the anterior pole of the oocyte and encodes a homedomain transcription factor that specifies the anterior of the developing embryo [
27,
28]. The
osk mRNA is transported to the posterior pole of the oocyte. Oskar is an RNA scaffold protein that (1) recruits the
nanos mRNA whose encoded protein controls the posterior segmentation of the embryo and (2) recruits the determinants necessary to specify the future germ cells at the posterior of the embryo [
29,
30,
31,
32,
33]. The
grk mRNA is transported close to the oocyte nucleus and relocalises with it when the nucleus migrates to an asymmetric position [
34]. The Grk protein is a TGF-alpha ortholog [
34]. Upon its local translation, it is secreted toward the nearby follicle cells, where it activates the EGF receptor (EGFr) and triggers several specific differentiation programs in those cells [
7].
-
During early oogenesis, before stage 6, the
grk mRNA is localized close to the nucleus hemisphere facing the posterior of the oocyte, and its translation leads to the activation of the EGFr in about 20 follicle cells adjacent to the oocyte. This induces a differentiation program necessary for building the posterior structures of the eggshell [
35,
36,
37,
38] and for the emission, in conjunction with the JAK-STAT signalling pathway, of a signal from those cells, that later triggers a repolarization of the MT network of the oocyte and the start of nucleus migration [
37,
38,
39] (
Figure 1C).
-
With the asymmetrical positioning of the nucleus to an antero-dorsal position, the
grk mRNA, still associated with the nucleus, relocalises in the vicinity of the antero-dorsal follicle cells where its translation triggers a second wave of EGFr activation in a gradient pattern (
Figure 1E). This enables the formation of two groups of follicle cells, which forms the dorsal appendages. Through a series of steps, the cells that lack EGFr activation because they do not receive the Grk signal, secrete the ligand triggering the formation of most of the different germ layers of the embryo via the activation of the Toll receptor [
7,
8]. A crucial step in this process is the asymmetrical positioning of the nucleus, which functions as a symmetry-breaking event for the formation of the dorsoventral axis of the eggshell and of the future embryo. Because the lateral follicle cells are all equivalent before this second wave of Grk signaling, and because there are no markers that predict where the nucleus would move within the oocyte, it is thought the nucleus can migrate towards any position representing the intersection between the anterior and lateral parts of the oocyte. Further support for this view comes from an elegant experiment showing in oocytes with two nuclei, the nuclei migrate to random positions with respect to each other [
40].
2. Steps in Nucleus Positioning during Oocyte Development
Nuclear positioning occurs between stages 5 and 7, with the nucleus assuming an asymmetric position at later stages [
18,
22,
23,
41]. A peculiarity of the oocyte nuclear migration is that it is a three-dimensional process. At stage 5, the oocyte can be assimilated to a truncated cone whose base is in contact with the apical part of the posterior follicular epithelium and the lateral and upper sides in contact with four nurse cells through ring canals and cell-cell contacts. At this stage, the oocyte has an anterior-posterior asymmetry, which is established after the oocyte is determined, such that the plasma membrane facing the posterior follicle cells constitutes the posterior pole of the oocyte and the plasma membrane in contact with the nurse cells the anterior side (
Figure 3A,B). At this stage, the dorsoventral polarity is not yet established, this being achieved with the asymmetric positioning of the nucleus as mentioned above. At stage 7, the shape of the oocyte develops into an asymmetric ellipse with the same characteristics for the anterior-posterior polarity. The nucleus is asymmetrically positioned at the intersection of the anterior and posterior plasma membranes, which corresponds to a ring at the edge of the anterior plasma membrane of the oocyte (
Figure 3A,B). Importantly, the nucleus can be located at any point on this circumference once its migration is complete as described above.
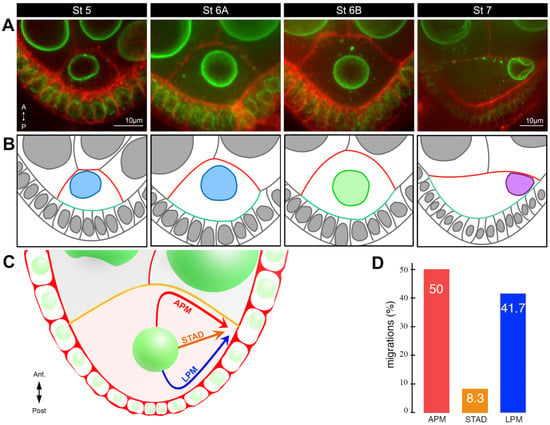
Figure 3. Characterisation of nucleus positioning before and during migration. (
A) Stage 5–7 egg chambers, expressing
Fs(2)Ket-GFP to label nuclei (green) and stained with Cellmask, a lipophilic dye to reveal plasma membranes (red). Representative examples of the different nuclear positions at stages 5, 6A, 6B, and 7 adapted from [
23]. The oocytes are oriented with anterior (
A) at the top and posterior (P) at the bottom. (
B) Schematic representations of the image above, with a color code illustrating the evolution of the oocyte shape (anterior plasma membrane in red, posterior plasma membrane in turquoise) and the position of the nucleus: anterior, pale blue; center, pale green; migrated to the intersection between anterior plasma membrane and the lateral plasma membrane, purple adapted from [
23]. (
C) Schematic illustration of the three alternative routes by which the nucleus migrates to the antero-dorsal cortex in the oocyte: along the anterior plasma membrane (APM), through the cytoplasm (STAD), or along the lateral plasma membrane (LPM) [
18]. (
D) Bar plots of the distribution of the three different migration paths taken by the nuclei [
18].
The positioning of the oocyte nucleus is a dynamic process during oocyte development. From the germarium to stage 5, the nucleus is positioned close to the anterior plasma membrane (
Figure 1A). From stage 5 onwards, the nucleus gradually assumes a central position until stage 6B before migration starts (
Figure 3A,B). This centration process relies on the clustering of centrosomes (see below) [
23]. This suggests the nucleus has to be centered in the oocyte to migrate. Migration is a relatively slow process taking about two hours for completion [
18]. Interestingly, it is a variable process that can be achieved in different ways [
18]. In most cases, a biphasic migration pattern was observed. The nucleus hits either the anterior or the lateral plasma membranes before sliding along them to reach its final destination. However, in rare cases (8%), the nucleus migrates directly to the antero-dorsal cortex and does not come into contact with the plasma membrane before its arrival (
Figure 3C,D). From stage 7 onward, the nucleus is maintained in its antero-dorsal position until the end of oogenesis and fertilisation. This anchoring is essential to ensure the correct establishment of the dorsoventral polarity of the egg chamber and of the future embryo. Importantly, it has been shown if an asymmetric position of the nucleus was not subsequently maintained, the dorsoventral axis was not determined correctly [
42]. Moreover, the antero-dorsal positioning of the nuclei, which is maintained until the end of oogenesis, is likely to be important for the formation of the zygote, as the sperm enters the oocyte from the antero-dorsal side through a channel, the micropyle. Its positioning is controlled during its morphogenesis by the TGF-alpha Grk, which is associated with the oocyte nucleus [
9,
43] (see below).
Oocyte Nucleus Positioning and Cytoskeleton
Initial studies evaluating the effect on oocyte development of MT-depolymerising drugs such as colchicine, reported a mispositioning of the oocyte nucleus, highlighting a critical role for MTs [
16,
44]. These results were further supported by the identification of mutations that affected the dynamics of MTs and impaired nuclear positioning, such as a mutation of the
Drosophila tubulin-binding cofactor B that enabled the assembly of the alpha tubulin and beta tubulin heterodimer complex [
45]. In contrast, although the nucleus is surrounded by a faint ring of actin at mid-oogenesis, latrunculin B, an actin depolymerising drug, at concentrations sufficient to abolish phalloidin-mediated actin detection and impair egg chamber morphogenesis, does not affect nuclear migration (Chemla and Guichet 2024 [
46]. Accordingly, a mutation affecting either Cappuccino, a formin, or Chickadee, a profilin [
47], both of which are involved in actin assembly in the oocyte, impair MT-based mRNA-associated transport in the oocyte without affecting nuclear positioning. However, it should be noted latrunculin B does not necessarily induce a full depolymerisation of the actin networks. In addition, the
capuccino and
chickadee alleles that affect oocyte polarity are not null alleles [
47], while a total loss of Chickadee activity prevents oocyte formation [
48]. It remains, therefore, to be determined whether actin plays a complementary role to the essential role of MTs.
Prior to its migration the nucleus oscillates around a central position [
18]. Given the complexity of the MT network, it was challenging to understand how MTs applied their forces on the nucleus. Depending on whether the MT reorganisation that leads to an inversion of the polarity of the MT networks after the reception of the back signal [
16,
22,
37,
38] occurs before or after nucleus migration, one can envision the nucleus is either pulled by an anterior network or pushed by a posterior network. Live imaging experiments associated with laser-mediated nanosurgery ablations of MTs have shown the nucleus was mainly pushed by the MTs to reach its final destination and the reorganisation of MT network did not occur prior to nucleus migration [
18,
22]. Furthermore, the nucleus itself is involved in the MT network reorganisation, as is illustrated by the detection of an abnormal MT organisation in oocytes in which the nucleus fails to migrate or is mispositioned [
16,
42]. In fact, several MT networks have been found to be required for the nuclear migration [
18].
This entry is adapted from the peer-reviewed paper 10.3390/cells13020201