Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Virology
Bovine leukemia virus (BLV) is the causative agent of enzootic bovine leukosis (EBL), which is the most significant neoplastic disease in cattle. EBL is often overlooked in daily breeding processes due to the absence of obvious clinical symptoms.
- bovine leukemia virus (BLV)
- genomic structure
- epidemiology
- transmission
1. Introduction
Bovine leukemia virus (BLV) is a retrovirus known to cause enzootic bovine leucosis (EBL) [1,2,3,4]. EBL has been successfully eradicated in most European countries [5]. However, recent investigations have shown an increase in infection rates in several countries, including Argentina, Brazil, Canada, Japan, and the United States [6,7,8,9]. The majority of BLV-infected cattle remain asymptomatic, harboring the virus in a latent state of infection, and are thus classified as carriers. As the disease progresses, around one-third of cattle infected with BLV exhibit a non-malignant proliferation of B lymphocytes, a condition referred to as persistent lymphocytosis (PL). After a long period of latent infection lasting 3 to 8 years, less than 5% of these cattle will eventually develop malignant B lymphocyte lymphoma [10,11].
Previous research suggested that lymphosarcoma caused by BLV primarily affected the economics of cattle farming. However, recent research has shown that BLV infection affects the immune system of cattle, even during the latency period, leading to varying degrees of immune suppression and direct economic losses. These losses include reduced milk production, increased susceptibility to infectious diseases (such as mastitis, skin infections, and hoof diseases), and decreased reproductive efficiency [12,13,14,15]. In the United States, where the BLV infection rate exceeds 40%, the annual losses due to reduced milk production have surpassed USD 500 million. Additionally, certain countries and regions have implemented strict import and export regulations for animals infected with BLV, resulting in various indirect economic losses [5,13,15,16,17,18]. As a result, the World Organisation for Animal Health (WOAH) has classified Enzootic Bovine Leukosis (EBL) as a disease that significantly impacts international trade [19].
In addition to its impact on cattle herds, studies have shown that unsterilized or uncooked dairy and beef products contain proviral nucleic acid fragments of BLV, which raises concerns about public health and safety [20]. While it has not been confirmed that BLV can cause human infections, the positive detection rate of BLV in samples from breast cancer patients is significantly higher compared to healthy breast control groups [21,22,23,24,25,26]. Moreover, there is a correlation between the consumption of beef, milk, and dairy products and the incidence of breast cancer in different countries and regions. For example, countries with lower consumption rates like India, Japan, South Korea, and China also exhibit lower incidences of breast cancer. Conversely, countries with higher consumption rates such as the United States, the United Kingdom, Australia, and Germany have a higher incidence of breast cancer [27,28]. However, some studies have not detected proviral DNA of BLV in breast cancer tissue samples [29,30,31].
2. Genomic Composition of BLV
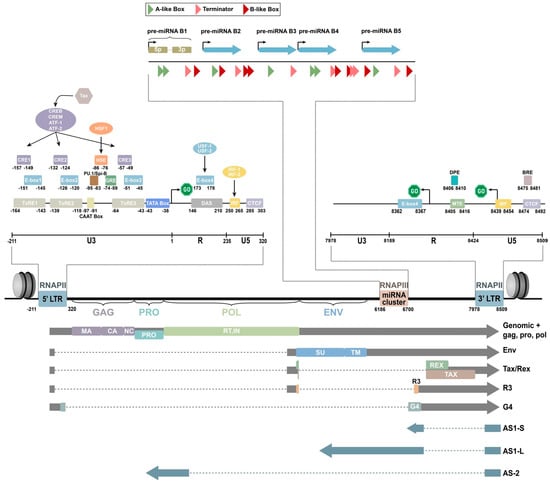
BLV is a single-stranded RNA-RT virus that exhibits a spherical or rod-shaped morphology, measuring approximately 60 to 125 nanometers in diameter. It is enveloped by a double-layered membrane structure and its nucleocapsid demonstrates icosahedral symmetry [32,33]. It contains a diploid single-stranded RNA genome. The viral genome of BLV consists of a diploid single-stranded RNA molecule spanning a total of 8714 nucleotides. This genome encompasses structural protein genes, enzyme coding genes, the pX region, and two identical, long, terminal repeat sequences positioned at both ends of the genome (Figure 1) [34,35].
Figure 1. Schematic representation of the BLV genome [35].
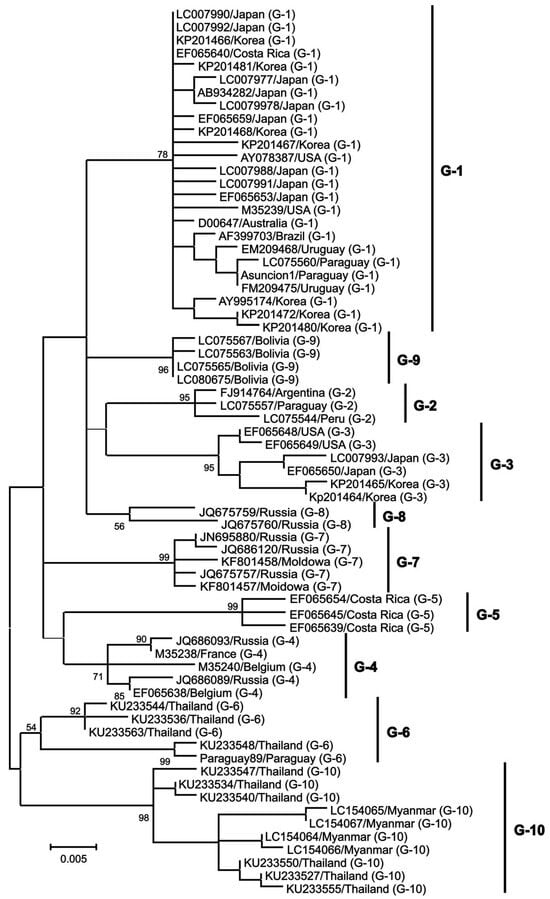
Structural proteins and enzyme coding genes, such as gag, pro, pol, and env, are crucial in the virus lifecycle, infectivity, and the production of infectious viral particles [36,37,38]. The gag gene is highly conserved and responsible for encoding three major non-glycosylated proteins: p12 nucleocapsid protein (which binds with viral genomic RNA), p15 matrix protein (which binds with viral genomic RNA and interacts with the lipid bilayer of the viral membrane), and p24 nucleocapsid protein (which serves as the main target of the host immune response) [37]. The pro gene encodes the viral proteinase p14, which is involved in the post-translation maturation of the virus [36]. The pol gene encodes reverse transcriptase and integrase, which play a role in the reverse transcription process and integration of proviral BLV DNA into the host genome [39]. The env gene encodes two glycosylated proteins: envelope protein gp51 and transmembrane protein gp30, which facilitate virus–cell fusion through interaction with cell membrane receptors [40,41]. The pX region, located between the env gene and 3’LTR, encodes other auxiliary regulatory or non-structural genes, including regulatory proteins Tax and Rex, as well as auxiliary proteins R3 and G4 [42,43,44,45,46,47,48]. Regulatory proteins are important in virus transcription regulation, inducing the malignant transformation of tumors and virus infection output, while R3 and G4 auxiliary proteins help maintain a high viral load [49]. The env gene also shows genetic polymorphism, which is beneficial for phylogenetic studies and classifying BLV isolates [40]. The nucleotide sequence and amino acid composition of the env gene are valuable genomic markers for studying the distribution of BLV and identifying different genotypes based on geographical origin [50]. Researchers have identified 10 BLV genotypes through sequencing and phylogenetic analysis of partial and full-length gp51 env genes. Genotype 1 is the most prevalent and commonly found in the United States, Japan, and South Korea [51,52,53]. South America has genotypes 1, 2, 3, 4, 5, 6, and 9 (with genotype 9 being particularly prevalent in Bolivia). Russia and Eastern Europe commonly have genotypes 4, 7, and 8 [54]. Genotype 10 is prevalent in China, Vietnam, Thailand, and Myanmar [51,55]. Genotype 4 is found in various countries in North and South America, Africa, and Asia [56,57]. Genotype 6 has been found in South American countries such as Argentina, Brazil, Peru, Paraguay, and Bolivia, as well as in Asian countries such as the Philippines, Thailand, and India [56,58,59,60]. Additionally, genotype 1 has been reported in Egypt [61]. Monitoring the emergence of new viral mutations in veterinary medicine and animal husbandry is important since each new genotype may exhibit unique characteristics in its interaction with the host organism and leukemogenesis is influenced by specific genotypes (Figure 2).
Figure 2. A maximum likelihood phylogenetic tree was constructed using partial BLV env sequences from various geographical locations worldwide, from [62].
Research indicates that 10 different microRNA (miRNA) can be transcribed under the control of RNA polymerase III in the genomic region spanning from the env gene to the R3 coding area [63]. RNA sequencing analysis has revealed that these miRNAs are highly expressed in primary sheep and cattle cells containing pre-leukemic and malignant B cells, accounting for 40% of the total cellular miRNA [64]. Additionally, elevated levels of BLV miRNA were found in the serum of infected animals, suggesting a potential paracrine role. These findings indicate the important role of BLV miRNAs in the retroviral cycle and/or disease progression, challenging the traditional belief that RNA viruses do not encode miRNAs. They also demonstrate that BLV miRNAs regulate various biological processes, including apoptosis, immunity, signal transduction, and tumorigenesis [65]. Notably, Blv-miR-B4-3p, which shares a seed sequence with the host miRNA miR-29a, is known for downregulating the tumor suppressor genes HMG-box transcription factor 1 (HBP1) and peroxidasin homologs (PXDN) in B-cell tumors [63,65,66]. In vitro studies have confirmed that blv-miR-B4-3p can directly downregulate these genes, although this has not been experimentally validated in cattle infected with BLV. Recently, Petersen et al. used RT-qPCR to analyze hbp1 and pxdn expression in cattle naturally infected with BLV and noted a significant downregulation of pxdn compared to uninfected animals [67]. However, a comprehensive analysis of targets for each mature BLV miRNA is still needed to fully understand the role of BLV in tumorigenesis.
This entry is adapted from the peer-reviewed paper 10.3390/ani14020297
This entry is offline, you can click here to edit this entry!