The human Dickkopf (DKK) family includes four main secreted proteins, DKK-1, DKK-2, DKK-3, and DKK-4, as well as the DKK-3 related protein soggy (Sgy-1 or DKKL1). These glycoproteins play crucial roles in various biological processes, and especially modulation of the Wnt signaling pathway. DKK-3 is distinct, with its multifaceted roles in development, stem cell differentiation and tissue homeostasis. Intriguingly, DKK-3 appears to have immunomodulatory functions and a complex role in cancer, acting as either a tumor suppressor or an oncogene, depending on the context. DKK-3 is a promising diagnostic and therapeutic target that can be modulated by epigenetic reactivation, gene therapy and DKK-3-blocking agents.
- Dickkopf (DKK) family
- Wnt signaling pathway
- DKK-3
- development
- immunomodulation
- cancer
1. Introduction
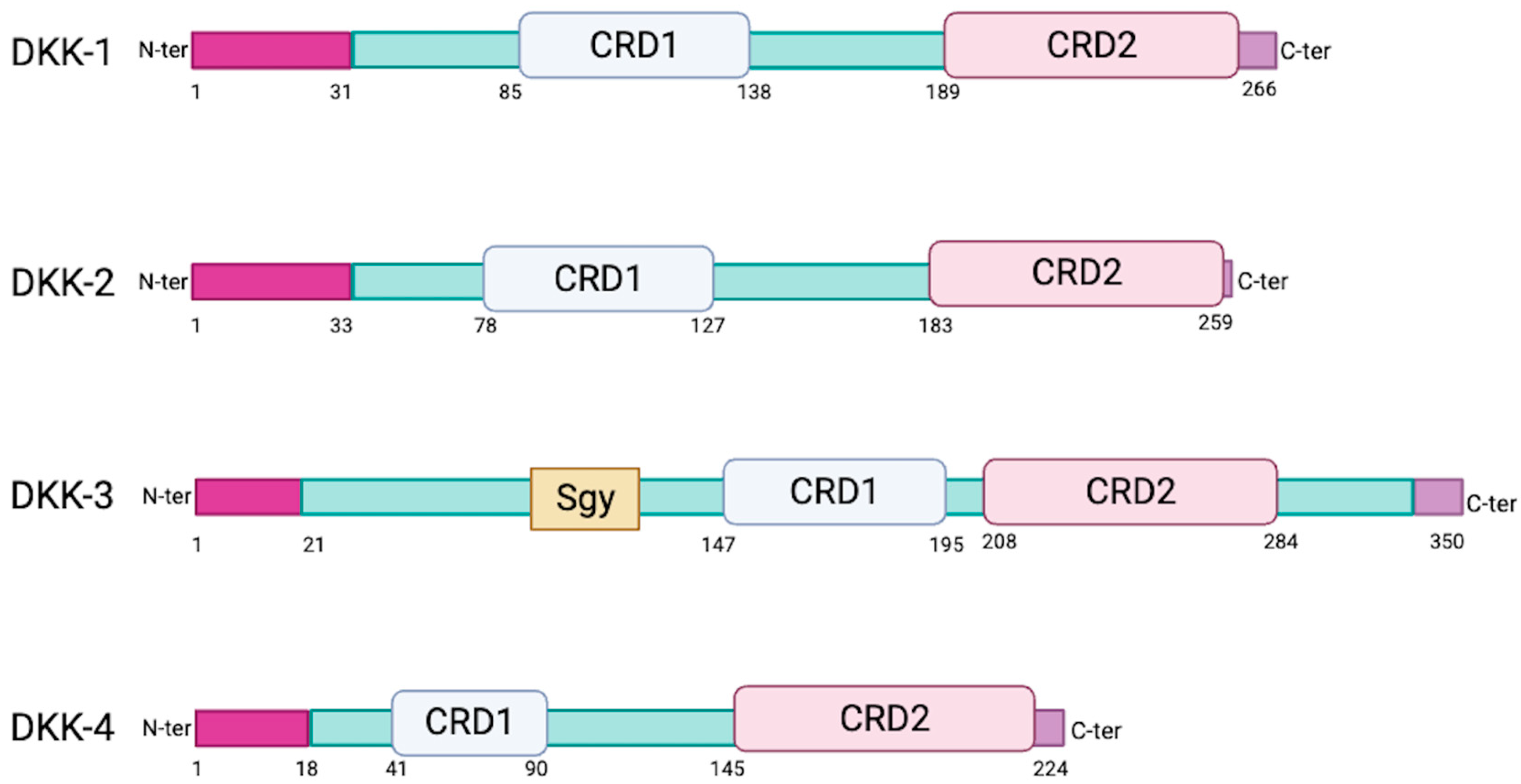
2. Sequence and Structural Properties of the DKK Family of Proteins
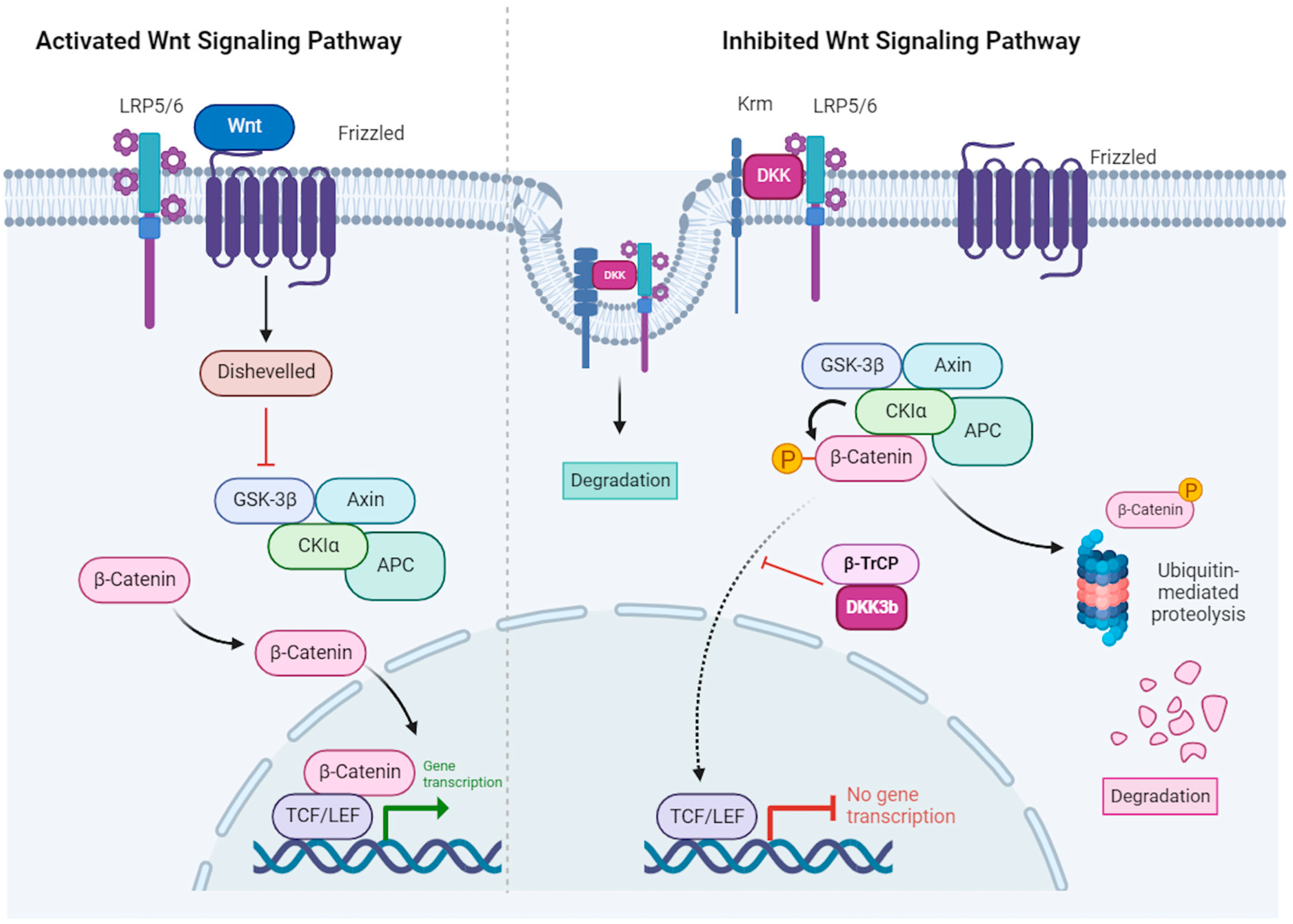
3. DKKs Modulate the Wnt Signaling Pathway through Binding to the LRP5/6 and Krm1/2 Co-Receptors
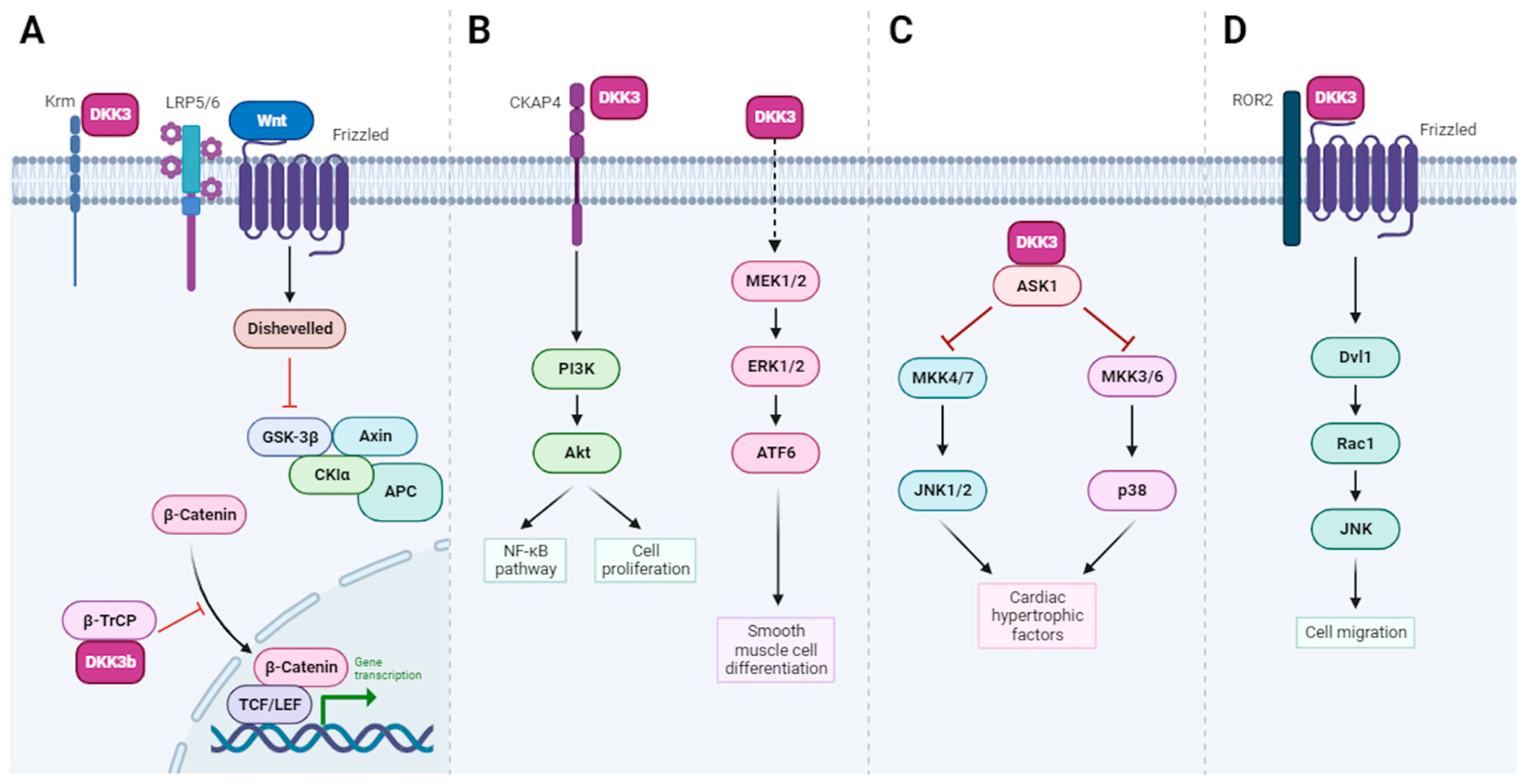
4. CKAP4 Is the Only Known Receptor for DKK-3
5. DKK-3 Has Diverse Biological Roles
| DKK-3 Function | Comment | Ref. | |
|---|---|---|---|
| Cell protection | Cell adhesion, migration and invasion | Secreted DKK-3 is part of the extracellular matrix and promotes cell adhesion, migration and invasion | [38] |
| Chondroprotection | DKK-3 prevents interleukin-1β/oncostatin-M driven proteoglycan loss of cartilage and is upregulated in osteoarthritis | [39] | |
| Protection against cardiac hypertrophy | DKK-3 protects the heart against pressure overload-induced cardiac remodeling through the inhibition of the ASK1-JNK/p38 signaling pathway | [40] | |
| Smooth muscle cell (SMC) differentiation | DKK-3 induces the expression of differentiated SMC markers (myocardin) through the activation of the ATF6 transcription factor | [41] | |
| Endothelium protection and repair | DKK-3−/− mice have more atherosclerotic lesions. DKK-3 stimulates endothelial cell migration through the Wnt/planar cell polarity signaling pathway | [42] | |
| Protection against neuronal death | DKK-3−/− ischemic mice have greater infarct size. DKK-3 prevents astrocyte death via the upregulation of VEGF expression and the activation of VEGFR2 | [43] | |
| Immune response and inflammation | B-cell fate and function | B1 cells have better proliferation and survival abilities whereas the development of B2 cells is impaired in DKK-3−/− mice, which also show altered immune and cytokine responses | [44] |
| Peripheral CD8 T-cell tolerance |
|
[45][46] | |
| Anti-tumor immune response regulation | High expression of DKK-3 in glioblastoma multiform correlates with poorer prognosis and decreased anti-tumor immunity. | [47] | |
| Regulation of inflammatory cytokines | DKK-3 reduces the intracerebral hemorrhage-related expression of TNF-α and IL-1β | [25] | |
| Differentiation of monocytes to DCs | Human CD14+ monocytes grown with recombinant DKK-3 differentiate into immature CD11c+, CD40+, CD86+, HLADR+ DCs | [48] | |
| Microglia polarization and reduction of neuroinflammation | DKK-3 inhibits the ASK-1/JNK/p38 pathway, favoring the transformation of type M1 to M2 microglia and the downregulation of TNF-α, IL-6 and IL-1β | [25] | |
| Tumor suppressor function | Downregulated expression in cancer cells | DKK-3 expression is lower in immortalized cell lines and 8 human tumor-derived cell lines | [49] |
| DKK-3 promoter is hypermethylated in cancer | The frequency of DKK-3 promoter hypermethylation varies from 26% (pleural mesothelia) to 61% (breast carcinoma). | [7][50] | |
| Suppression of cell proliferation | The overexpression of DKK-3 in hepatoma and cervical cancer cells diminishes cell proliferation in vitro | [51][52] | |
| Response to chemotherapy |
|
[15][53] | |
| Serum DKK-3 is a prognosis biomarker | DKK-3 levels are negatively correlated with the number of CD133+ circulating tumor cells in ovarian cancer | [54] | |
| Regulation of the tumor immune microenvironment | DKK-3 expression correlates with infiltrating CD8+ lymphocytes and macrophages in HPV-positive HNSCC and stimulates NF-κB signaling in macrophages | [55] | |
| Oncogenic function | Stimulation of cell proliferation, migration and tumor growth | The expression of DKK-3 is high in HNSCC, esophageal and pancreatic cell lines, where it stimulates cell proliferation and migration in vitro. In xenograft models, DKK-3 supports tumor growth. | [10][56][57][58] |
| Correlation with tumor progression | DKK-3 expression was found to increase from normal oral mucosa in dysplasia and squamous cell carcinoma and to correlate with the tissue proliferative index (Ki67) | [59] | |
| DKK-3 has pro-tumorigenic function in tumor stroma |
|
[26][60] | |
| DKK-3 impairs anti-cancer immunity |
|
[29][30][31][47][61][62] |
6. DKK-3 Modulates the Immune System
7. DKK-3 Has a Dual Role in Cancer, as Either a Tumor Suppressor or an Oncogene
7.1. DKK-3 as a Tumor Suppressor
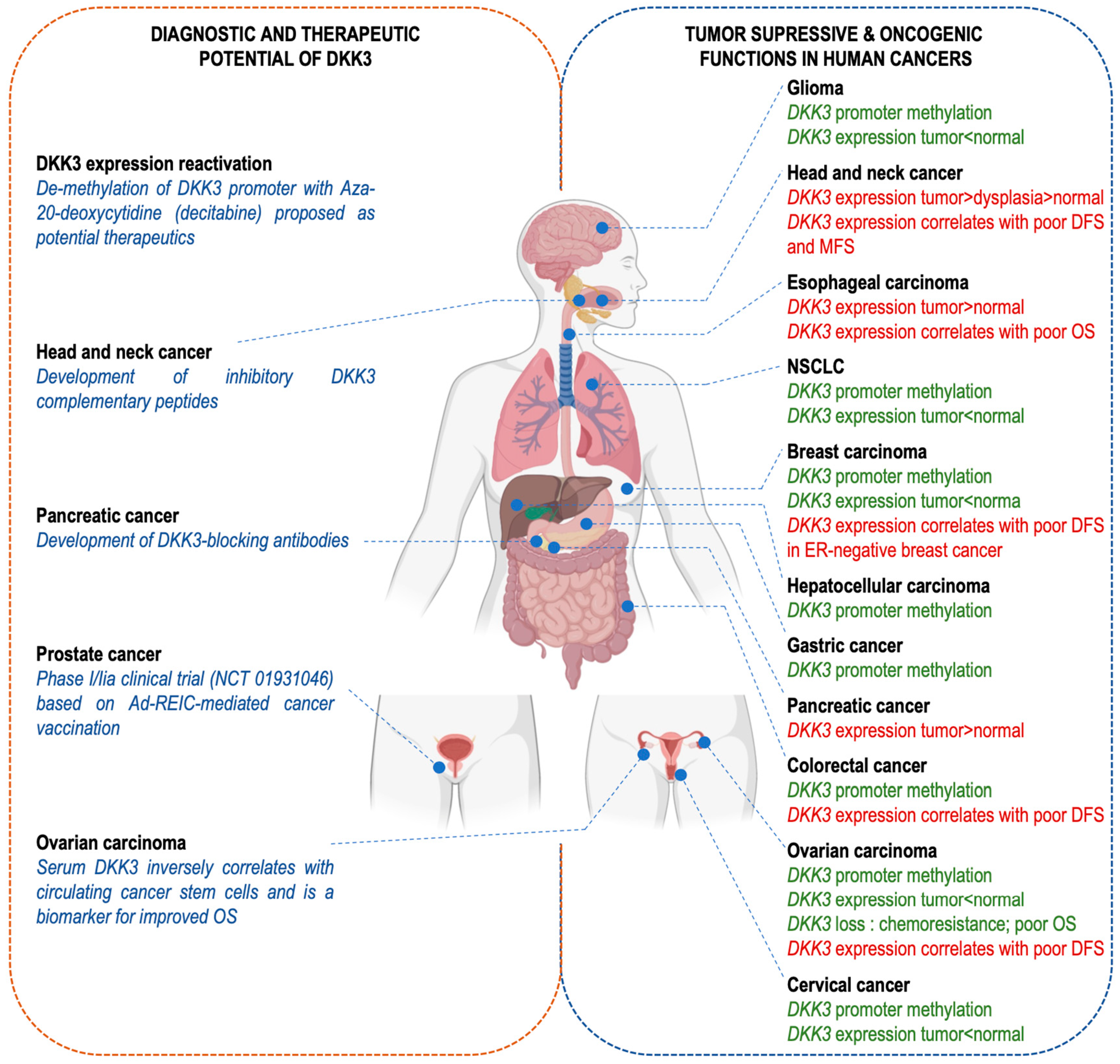
7.2. DKK-3 as an Oncogene
8. DKK-3 as a Tool for Cancer Therapy
This entry is adapted from the peer-reviewed paper 10.3390/cells13010075
References
- Mannino, G.; Russo, C.; Maugeri, G.; Musumeci, G.; Vicario, N.; Tibullo, D.; Giuffrida, R.; Parenti, R.; Lo Furno, D. Adult Stem Cell Niches for Tissue Homeostasis. J. Cell Physiol. 2022, 237, 239–257.
- Aiello, N.M.; Stanger, B.Z. Echoes of the Embryo: Using the Developmental Biology Toolkit to Study Cancer. Dis. Models Mech. 2016, 9, 105–114.
- Zhang, Y.; Wang, X. Targeting the Wnt/β-Catenin Signaling Pathway in Cancer. J. Hematol. Oncol. 2020, 13, 165.
- Kikuchi, A.; Matsumoto, S.; Sada, R. Dickkopf Signaling, beyond Wnt-Mediated Biology. Semin. Cell Dev. Biol. 2022, 125, 55–65.
- Giralt, I.; Gallo-Oller, G.; Navarro, N.; Zarzosa, P.; Pons, G.; Magdaleno, A.; Segura, M.F.; Sánchez de Toledo, J.; Moreno, L.; Gallego, S.; et al. Dickkopf Proteins and Their Role in Cancer: A Family of Wnt Antagonists with a Dual Role. Pharmaceuticals 2021, 14, 810.
- Kaneko, K.J.; DePamphilis, M.L. Soggy, a Spermatocyte-Specific Gene, Lies 3.8 Kb Upstream of and Antipodal to TEAD-2, a Transcription Factor Expressed at the Beginning of Mouse Development. Nucleic Acids Res. 2000, 28, 3982–3990.
- Veeck, J.; Dahl, E. Targeting the Wnt Pathway in Cancer: The Emerging Role of Dickkopf-3. Biochim. Et Biophys. Acta (BBA)-Rev. Cancer 2012, 1825, 18–28.
- Krupnik, V.E.; Sharp, J.D.; Jiang, C.; Robison, K.; Chickering, T.W.; Amaravadi, L.; Brown, D.E.; Guyot, D.; Mays, G.; Leiby, K.; et al. Functional and Structural Diversity of the Human Dickkopf Gene Family. Gene 1999, 238, 301–313.
- Niehrs, C. Function and Biological Roles of the Dickkopf Family of Wnt Modulators. Oncogene 2006, 25, 7469–7481.
- Kajiwara, C.; Fumoto, K.; Kimura, H.; Nojima, S.; Asano, K.; Odagiri, K.; Yamasaki, M.; Hikita, H.; Takehara, T.; Doki, Y.; et al. P63-Dependent Dickkopf3 Expression Promotes Esophageal Cancer Cell Proliferation via CKAP4. Cancer Res. 2018, 78, 6107–6120.
- Kawano, Y.; Kypta, R. Secreted Antagonists of the Wnt Signalling Pathway. J. Cell Sci. 2003, 116, 2627–2634.
- Hayat, R.; Manzoor, M.; Hussain, A. Wnt Signaling Pathway: A Comprehensive Review. Cell Biol. Int. 2022, 46, 863–877.
- Aravind, L.; Koonin, E.V. A Colipase Fold in the Carboxy-Terminal Domain of the Wnt Antagonists—The Dickkopfs. Curr. Biol. 1998, 8, R477–R479.
- Kohn, A.D.; Moon, R.T. Wnt and Calcium Signaling: Beta-Catenin-Independent Pathways. Cell Calcium. 2005, 38, 439–446.
- Wang, Z.; Ma, L.-J.; Kang, Y.; Li, X.; Zhang, X.-J. Dickkopf-3 (Dkk3) Induces Apoptosis in Cisplatin-Resistant Lung Adenocarcinoma Cells via the Wnt/β-Catenin Pathway. Oncol. Rep. 2015, 33, 1097–1106.
- Duchartre, Y.; Kim, Y.-M.; Kahn, M. The Wnt Signaling Pathway in Cancer. Crit. Rev. Oncol. Hematol. 2016, 99, 141–149.
- Schunk, S.J.; Floege, J.; Fliser, D.; Speer, T. WNT–β-catenin signalling—A versatile player in kidney injury and repair. Nat. Rev. Nephrol. 2021, 17, 172–184.
- MacDonald, B.T.; He, X. Frizzled and LRP5/6 Receptors for Wnt/β-Catenin Signaling. Cold Spring Harb. Perspect. Biol. 2012, 4, a007880.
- Mao, B.; Wu, W.; Li, Y.; Hoppe, D.; Stannek, P.; Glinka, A.; Niehrs, C. LDL-Receptor-Related Protein 6 Is a Receptor for Dickkopf Proteins. Nature 2001, 411, 321–325.
- Mao, B.; Wu, W.; Davidson, G.; Marhold, J.; Li, M.; Mechler, B.M.; Delius, H.; Hoppe, D.; Stannek, P.; Walter, C.; et al. Kremen Proteins Are Dickkopf Receptors That Regulate Wnt/b-Catenin Signalling. Nature 2002, 417, 664–667.
- Nakamura, T.; Aoki, S.; Kitajima, K.; Takahashi, T.; Matsumoto, K.; Nakamura, T. Molecular Cloning and Characterization of Kremen, a Novel Kringle-Containing Transmembrane Protein. Biochim. Et Biophys. Acta (BBA)-Gene Struct. Expr. 2001, 1518, 63–72.
- Mao, B.; Niehrs, C. Kremen2 Modulates Dickkopf2 Activity during Wnt/lRP6 Signaling. Gene 2003, 302, 179–183.
- Mohammadpour, H.; Khalili, S.; Hashemi, Z.S. Kremen Is beyond a Subsidiary Co-Receptor of Wnt Signaling: An in Silico Validation. Turk. J. Biol. 2015, 39, 501–510.
- Mohammadpour, H.; Pourfathollah, A.A.; Nikougoftar Zarif, M.; Khalili, S. Key Role of Dkk3 Protein in Inhibition of Cancer Cell Proliferation: An In Silico Identification. J. Theor. Biol. 2016, 393, 98–104.
- Xu, Y.; Nowrangi, D.; Liang, H.; Wang, T.; Yu, L.; Lu, T.; Lu, Z.; Zhang, J.H.; Luo, B.; Tang, J. DKK3 Attenuates JNK and AP-1 Induced Inflammation via Kremen-1 and DVL-1 in Mice Following Intracerebral Hemorrhage. J. Neuroinflamm. 2020, 17, 130.
- Ferrari, N.; Ranftl, R.; Chicherova, I.; Slaven, N.D.; Moeendarbary, E.; Farrugia, A.J.; Lam, M.; Semiannikova, M.; Westergaard, M.C.W.; Tchou, J.; et al. Dickkopf-3 Links HSF1 and YAP/TAZ Signalling to Control Aggressive Behaviours in Cancer-Associated Fibroblasts. Nat. Commun. 2019, 10, 130.
- Leonard, D.M.; Stachelek, S.J.; Safran, M.; Farwell, A.P.; Kowalik, T.F.; Leonard, J.L. Cloning, Expression, and Functional Characterization of the Substrate Binding Subunit of Rat Type II Iodothyronine 5′-Deiodinase. J. Biol. Chem. 2000, 275, 25194–25201.
- Stachelek, S.J.; Tuft, R.A.; Lifschitz, L.M.; Leonard, D.M.; Farwell, A.P.; Leonard, J.L. Real-Time Visualization of Processive Myosin 5a-Mediated Vesicle Movement in Living Astrocytes * 210. J. Biol. Chem. 2001, 276, 35652–35659.
- Katase, N.; Nagano, K.; Fujita, S. DKK3 Expression and Function in Head and Neck Squamous Cell Carcinoma and Other Cancers. J. Oral Biosci. 2020, 62, 9–15.
- Leonard, J.L.; Leonard, D.M.; Wolfe, S.A.; Liu, J.; Rivera, J.; Yang, M.; Leonard, R.T.; Johnson, J.P.S.; Kumar, P.; Liebmann, K.L.; et al. The Dkk3 Gene Encodes a Vital Intracellular Regulator of Cell Proliferation. PLoS ONE 2017, 12, e0181724.
- Kimura, H.; Fumoto, K.; Shojima, K.; Nojima, S.; Osugi, Y.; Tomihara, H.; Eguchi, H.; Shintani, Y.; Endo, H.; Inoue, M.; et al. CKAP4 Is a Dickkopf1 Receptor and Is Involved in Tumor Progression. J. Clin. Investig. 2016, 126, 2689–2705.
- Sandoz, P.A.; Denhardt-Eriksson, R.A.; Abrami, L.; Abriata, L.A.; Spreemann, G.; Maclachlan, C.; Ho, S.; Kunz, B.; Hess, K.; Knott, G.; et al. Dynamics of CLIMP-63 S-Acylation Control ER Morphology. Nat. Commun. 2023, 14, 264.
- Kikuchi, A.; Fumoto, K.; Kimura, H. The Dickkopf1-Cytoskeleton-Associated Protein 4 Axis Creates a Novel Signalling Pathway and May Represent a Molecular Target for Cancer Therapy. Br. J. Pharmacol. 2017, 174, 4651–4665.
- Bhavanasi, D.; Speer, K.F.; Klein, P.S. CKAP4 Is Identified as a Receptor for Dickkopf in Cancer Cells. J. Clin. Investig. 2016, 126, 2419–2421.
- del Barrantes, I.B.; Montero-Pedrazuela, A.; Guadaño-Ferraz, A.; Obregon, M.-J.; Martinez de Mena, R.; Gailus-Durner, V.; Fuchs, H.; Franz, T.J.; Kalaydjiev, S.; Klempt, M.; et al. Generation and Characterization of Dickkopf3 Mutant Mice. Mol. Cell Biol. 2006, 26, 2317–2326.
- Bertozzi, A.; Wu, C.-C.; Hans, S.; Brand, M.; Weidinger, G. Wnt/β-Catenin Signaling Acts Cell-Autonomously to Promote Cardiomyocyte Regeneration in the Zebrafish Heart. Dev. Biol. 2022, 481, 226–237.
- Singh, S.P.; Chawla, P.; Hnatiuk, A.; Kamel, M.; Silva, L.D.; Spanjaard, B.; Eski, S.E.; Janjuha, S.; Olivares-Chauvet, P.; Kayisoglu, O.; et al. A Single-Cell Atlas of de Novo β-Cell Regeneration Reveals the Contribution of Hybrid β/δ-Cells to Diabetes Recovery in Zebrafish. Development 2022, 149, dev199853.
- Kano, J.; Wang, H.; Zhang, H.; Noguchi, M. Roles of DKK3 in Cellular Adhesion, Motility, and Invasion through Extracellular Interaction with TGFBI. FEBS J. 2022, 289, 6385–6399.
- Snelling, S.J.B.; Davidson, R.K.; Swingler, T.E.; Le, L.T.T.; Barter, M.J.; Culley, K.L.; Price, A.; Carr, A.J.; Clark, I.M. Dickkopf-3 Is Upregulated in Osteoarthritis and Has a Chondroprotective Role. Osteoarthr. Cartil. 2016, 24, 883–891.
- Zhang, Y.; Liu, Y.; Zhu, X.-H.; Zhang, X.-D.; Jiang, D.-S.; Bian, Z.-Y.; Zhang, X.-F.; Chen, K.; Wei, X.; Gao, L.; et al. Dickkopf-3 Attenuates Pressure Overload-Induced Cardiac Remodelling. Cardiovasc. Res. 2014, 102, 35–45.
- Wang, X.; Karamariti, E.; Simpson, R.; Wang, W.; Xu, Q. Dickkopf Homolog 3 Induces Stem Cell Differentiation into Smooth Muscle Lineage via ATF6 Signalling. J. Biol. Chem. 2015, 290, 19844–19852.
- Yu, B.; Kiechl, S.; Qi, D.; Wang, X.; Song, Y.; Weger, S.; Mayr, A.; Le Bras, A.; Karamariti, E.; Zhang, Z.; et al. A Cytokine-like Protein Dickkopf-Related Protein 3 Is Atheroprotective. Circulation 2017, 136, 1022–1036.
- Busceti, C.L.; Di Menna, L.; Bianchi, F.; Mastroiacovo, F.; Di Pietro, P.; Traficante, A.; Bozza, G.; Niehrs, C.; Battaglia, G.; Bruno, V.; et al. Dickkopf-3 Causes Neuroprotection by Inducing Vascular Endothelial Growth Factor. Front. Cell Neurosci. 2018, 12, 292.
- Ludwig, J.; Federico, G.; Prokosch, S.; Küblbeck, G.; Schmitt, S.; Klevenz, A.; Gröne, H.-J.; Nitschke, L.; Arnold, B. Dickkopf-3 Acts as a Modulator of B Cell Fate and Function. J. Immunol. 2015, 194, 2624–2634.
- Papatriantafyllou, M.; Moldenhauer, G.; Ludwig, J.; Tafuri, A.; Garbi, N.; Hollmann, G.; Küblbeck, G.; Klevenz, A.; Schmitt, S.; Pougialis, G.; et al. Dickkopf-3, an Immune Modulator in Peripheral CD8 T-Cell Tolerance. Proc. Natl. Acad. Sci. USA 2012, 109, 1631–1636.
- Meister, M.; Papatriantafyllou, M.; Nordström, V.; Kumar, V.; Ludwig, J.; Lui, K.O.; Boyd, A.S.; Popovic, Z.V.; Fleming, T.H.; Moldenhauer, G.; et al. Dickkopf-3, a Tissue-Derived Modulator of Local T-Cell Responses. Front. Immunol. 2015, 6, 78.
- Han, M.-H.; Min, K.-W.; Noh, Y.-K.; Kim, J.M.; Cheong, J.H.; Ryu, J.I.; Won, Y.D.; Koh, S.-H.; Myung, J.K.; Park, J.Y.; et al. High DKK3 Expression Related to Immunosuppression Was Associated with Poor Prognosis in Glioblastoma: Machine Learning Approach. Cancer Immunol. Immunother. 2022, 71, 3013–3027.
- Watanabe, M.; Kashiwakura, Y.; Huang, P.; Ochiai, K.; Futami, J.; Li, S.-A.; Takaoka, M.; Nasu, Y.; Sakaguchi, M.; Huh, N.-H.; et al. Immunological Aspects of REIC/Dkk-3 in Monocyte Differentiation and Tumor Regression. Int. J. Oncol. 2009, 34, 657–663.
- Tsuji, T.; Miyazaki, M.; Sakaguchi, M.; Inoue, Y.; Namba, M. A REIC Gene Shows Down-Regulation in Human Immortalized Cells and Human Tumor-Derived Cell Lines. Biochem. Biophys. Res. Commun. 2000, 268, 20–24.
- Hayashi, T.; Asano, H.; Toyooka, S.; Tsukuda, K.; Soh, J.; Shien, T.; Taira, N.; Maki, Y.; Tanaka, N.; Doihara, H.; et al. DNA Methylation Status of REIC/Dkk-3 Gene in Human Malignancies. J. Cancer Res. Clin. Oncol. 2012, 138, 799–809.
- Hsieh, S.-Y.; Hsieh, P.-S.; Chiu, C.-T.; Chen, W.-Y. Dickkopf-3/REIC Functions as a Suppressor Gene of Tumor Growth. Oncogene 2004, 23, 9183–9189.
- Lee, E.-J.; Jo, M.; Rho, S.B.; Park, K.; Yoo, Y.-N.; Park, J.; Chae, M.; Zhang, W.; Lee, J.-H. Dkk3, Downregulated in Cervical Cancer, Functions as a Negative Regulator of β-Catenin. Int. J. Cancer 2009, 124, 287–297.
- Nguyen, Q.T.T.; Park, H.S.; Lee, T.J.; Choi, K.-M.; Park, J.Y.; Kim, D.; Kim, J.H.; Park, J.; Lee, E.-J. DKK3, Downregulated in Invasive Epithelial Ovarian Cancer, Is Associated with Chemoresistance and Enhanced Paclitaxel Susceptibility via Inhibition of the β-Catenin-P-Glycoprotein Signaling Pathway. Cancers 2022, 14, 924.
- Nie, X.-C.; He, F.; Lan, C.; Niu, J.-M.; Xia, P. Combined Serum DKK3 and Circulating CD133 Cells as Prognostic Biomarkers for Ovarian Cancer Patients. OncoTargets Ther. 2021, 14, 427–434.
- Mourtada, J.; Lony, C.; Nicol, A.; De Azevedo, J.; Bour, C.; Macabre, C.; Roncarati, P.; Ledrappier, S.; Schultz, P.; Borel, C.; et al. A Novel ΔNp63-Dependent Immune Mechanism Improves Prognosis of HPV-Related Head and Neck Cancer. Front. Immunol. 2023, 14, 1264093.
- Katase, N.; Nishimatsu, S.-I.; Yamauchi, A.; Yamamura, M.; Terada, K.; Itadani, M.; Okada, N.; Hassan, N.M.M.; Nagatsuka, H.; Ikeda, T.; et al. DKK3 Overexpression Increases the Malignant Properties of Head and Neck Squamous Cell Carcinoma Cells. Oncol. Res. 2018, 26, 45–58.
- Zhou, L.; Husted, H.; Moore, T.; Lu, M.; Deng, D.; Liu, Y.; Ramachandran, V.; Arumugam, T.; Niehrs, C.; Wang, H.; et al. Suppression of Stromal-Derived Dickkopf-3 (DKK3) Inhibits Tumor Progression and Prolongs Survival in Pancreatic Ductal Adenocarcinoma. Sci. Transl. Med. 2018, 10, eaat3487.
- Katase, N.; Nishimatsu, S.-I.; Yamauchi, A.; Yamamura, M.; Fujita, S. DKK3 Knockdown Confers Negative Effects on the Malignant Potency of Head and Neck Squamous Cell Carcinoma Cells via the PI3K/Akt and MAPK Signaling Pathways. Int. J. Oncol. 2019, 54, 1021–1032.
- Fujii, M.; Katase, N.; Lefeuvre, M.; Gunduz, M.; Buery, R.R.; Tamamura, R.; Tsujigiwa, H.; Nagatsuka, H. Dickkopf (Dkk)-3 and β-Catenin Expressions Increased in the Transition from Normal Oral Mucosal to Oral Squamous Cell Carcinoma. J. Mol. Hist. 2011, 42, 499–504.
- Zenzmaier, C.; Sampson, N.; Plas, E.; Berger, P. Dickkopf-related Protein 3 Promotes Pathogenic Stromal Remodeling in Benign Prostatic Hyperplasia and Prostate Cancer. Prostate 2013, 73, 1441–1452.
- Han, M.-H.; Baek, J.M.; Min, K.-W.; Cheong, J.H.; Ryu, J.I.; Won, Y.D.; Kwon, M.J.; Koh, S.-H. DKK3 Expression Is Associated with Immunosuppression and Poor Prognosis in Glioblastoma, in Contrast to Lower-Grade Gliomas. BMC Neurol. 2023, 23, 183.
- Lu, K.-H.; Tounsi, A.; Shridhar, N.; Küblbeck, G.; Klevenz, A.; Prokosch, S.; Bald, T.; Tüting, T.; Arnold, B. Dickkopf-3 Contributes to the Regulation of Anti-Tumor Immune Responses by Mesenchymal Stem Cells. Front. Immunol. 2015, 6, 645.
- Akazawa, H.; Komuro, I. Dickkopf-3: A Stubborn Protector of Cardiac Hypertrophy. Cardiovasc. Res. 2014, 102, 6–8.
- Zhang, L.-Q.; Gao, S.-J.; Sun, J.; Li, D.-Y.; Wu, J.-Y.; Song, F.-H.; Liu, D.-Q.; Zhou, Y.-Q.; Mei, W. DKK3 Ameliorates Neuropathic Pain via Inhibiting ASK-1/JNK/p-38-Mediated Microglia Polarization and Neuroinflammation. J. Neuroinflamm. 2022, 19, 129.
- Veeck, J.; Bektas, N.; Hartmann, A.; Kristiansen, G.; Heindrichs, U.; Knüchel, R.; Dahl, E. Wnt Signalling in Human Breast Cancer: Expression of the Putative Wnt Inhibitor Dickkopf-3 (DKK3) Is Frequently Suppressed by Promoter Hypermethylation in Mammary Tumours. Breast Cancer Res. 2008, 10, R82.
- Kumon, H.; Ariyoshi, Y.; Sasaki, K.; Sadahira, T.; Araki, M.; Ebara, S.; Yanai, H.; Watanabe, M.; Nasu, Y. Adenovirus Vector Carrying REIC/DKK-3 Gene: Neoadjuvant Intraprostatic Injection for High-Risk Localized Prostate Cancer Undergoing Radical Prostatectomy. Cancer Gene Ther. 2016, 23, 400–409.
- Oyama, A.; Shiraha, H.; Uchida, D.; Iwamuro, M.; Kato, H.; Takaki, A.; Ikeda, F.; Onishi, H.; Yasunaka, T.; Takeuchi, Y.; et al. A Phase I/Ib Trial of Ad-REIC in Liver Cancer: Study Protocol. Future Oncol. 2019, 15, 3547–3554.
- Romero, D.; Kypta, R. Dickkopf-3 Function in the Prostate. Bioarchitecture 2013, 3, 42–44.
- Wang, Z.; Lin, L.; Thomas, D.G.; Nadal, E.; Chang, A.C.; Beer, D.G.; Lin, J. The Role of Dickkopf-3 Overexpression in Esophageal Adenocarcinoma. J. Thorac. Cardiovasc. Surg. 2015, 150, 377–385.
- Katase, N.; Nishimatsu, S.-I.; Yamauchi, A.; Okano, S.; Fujita, S. DKK3 Expression Is Correlated with Poorer Prognosis in Head and Neck Squamous Cell Carcinoma: A Bioinformatics Study Based on the TCGA Database. J. Oral. Biosci. 2023, 65, 334–346.
- Hamzehzadeh, L.; Caraglia, M.; Atkin, S.L.; Sahebkar, A. Dickkopf Homolog 3 (DKK3): A Candidate for Detection and Treatment of Cancers? J. Cell. Physiol. 2018, 233, 4595–4605.
- Mehdipour, P.; Murphy, T.; De Carvalho, D.D. The Role of DNA-Demethylating Agents in Cancer Therapy. Pharmacol. Ther. 2020, 205, 107416.
- Watanabe, M.; Nasu, Y.; Kumon, H. Adenovirus-Mediated REIC/Dkk-3 Gene Therapy: Development of an Autologous Cancer Vaccination Therapy (Review). Oncol. Lett. 2014, 7, 595–601.
- Katase, N.; Lefeuvre, M.; Gunduz, M.; Gunduz, E.; Beder, L.B.; Grenman, R.; Fujii, M.; Tamamu, R.; Tsujigiwa, H.; Nagatsuka, H. Absence of Dickkopf (Dkk)-3 Protein Expression Is Correlated with Longer Disease-Free Survival and Lower Incidence of Metastasis in Head and Neck Squamous Cell Carcinoma. Oncol. Lett. 2012, 3, 273–280.
- Zenzmaier, C.; Hermann, M.; Hengster, P.; Berger, P. Dickkopf-3 Maintains the PANC-1 Human Pancreatic Tumor Cells in a Dedifferentiated State. Int. J. Oncol. 2012, 40, 40–46.
- Katase, N.; Nishimatsu, S.; Yamauchi, A.; Okano, S.; Fujita, S. Establishment of Anti-DKK3 Peptide for the Cancer Control in Head and Neck Squamous Cell Carcinoma (HNSCC). Cancer Cell Int. 2022, 22, 352.
- Marusyk, A.; Janiszewska, M.; Polyak, K. Intratumor Heterogeneity: The Rosetta Stone of Therapy Resistance. Cancer. Cell 2020, 37, 471–484.