Neurodegenerative diseases are a diverse group of neurological disorders that negatively impact the lives of millions of people across the world through the progressive loss of neurons [
1]. Among the neurodegenerative diseases, Alzheimer’s disease (AD) is linked to the majority of dementia cases. AD was initially reported 100 years ago by a German psychiatrist named Alois Alzheimer when he observed fibrillary structures within the brains of patients that came to him with progressive cognitive dysfunction [
2]. AD, along with other aging diseases, results in progressive lifestyle and cognitive decline, profoundly impacting the quality of life and functional independence of about 55 million people worldwide [
3]. The development of these diseases is attributed to the aggregation of misfolded proteins, which leads to progressive neuronal loss and subsequent dysfunction [
4]. These aggregates are insoluble fibrils characterized by a β-pleated sheet usually arranged in an antiparallel configuration [
5,
6]. In AD, β-amyloid (Aβ) aggregates into different oligomeric forms and amyloid fibrils. These fibrils are a major part of amyloid plaques in the brains of AD patients [
7]. Based on the involvement of Aβ aggregates in AD pathogenesis, they serve as a promising target for AD therapy [
8].
The formation of protein aggregates is the result of failed protein quality control machinery, the purpose of which is to refold or degrade misfolded proteins [
9]. A main component of the body’s quality control machinery is chaperone proteins. Molecular chaperones play a vital role in the body since they are the regulators of protein conformational states [
9,
10,
11]. Hence, chaperone proteins and targeting medicinal agents pose great potential for therapeutic intervention for AD and other related neurodegenerative diseases. The major molecular chaperone heat shock proteins (Hsps) are a family of stress proteins that play an important role in protein processing [
12,
13,
14]. Heat shock protein 90 (Hsp90) is a crucial molecular chaperone that is responsible for many mechanistic and therapeutic pathways in various diseases. The first report by Yoo et al. highlighted the significance of the deranged expression of HSPs and various other chaperones in AD cases [
15]. Furthermore, Hsp90 targeting compound 17-AAG has been shown to attenuate Aβ toxicity and prevent memory loss in AD models [
16]. Recent reports suggest that the Dihydropyridine derivative LA1011 compound’s binding to Hsp90 in pathological condition leads to heat shock response and neuroprotection [
17,
18].
The latest developments in AD research showed that the modulation of Hsp90, the BRICHOS domain, and several other prominent molecular chaperone proteins play a critical role in the regulation of Aβ processing. The name BRICHOS comes from BRI2, CHOndromodulin-I and Surfactant protein C. It is composed of 100 amino acid residues and has various fascinating properties including alleviating Aβ fibrillation and aggregation. Proteins that have this BRICHOS domain have been observed to have a diverse assortment of phenotypes including dementia, respiratory distress, chondrosarcoma, and stomach cancer [
20]. This domain has been seen in 12 different protein families up until this point. All BRICHOS domain containing proteins and Buxbaum et al. provided here [
21]. These proteins harbor similar predicted secondary structures [
21]. The BRICHOS domain has also been linked to lung fibrosis, dementia, and cancer [
22,
23,
24]. Artificial chaperones are chaperone–mimetic molecules that selectively associate with client substrates for proper folding and processing. These chaperones have been developed for clinical applications [
25].
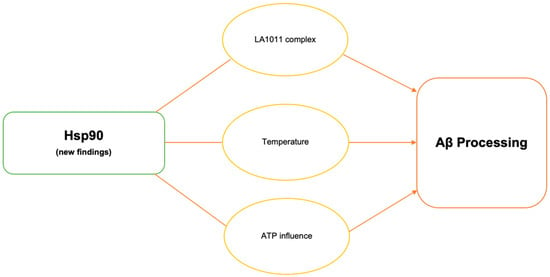
2. Hsp90
2.1. Hsp90–LA1011 Complex
Hsp90 is capable of both solubilizing and suppressing protein aggregates and is also the most prevalent chaperone in mechanistic pathways. Several reports suggested that Hsp90-targeting compounds represent great potential for the development of AD therapy. Recently, Hsp co-inducer 1,4-dihydropyridine derivative LA1011 was shown to have a neuroprotective effect [
17]. One of these major discoveries lies in LA1011 complex formation and explores how its structure works with various co-chaperones of Hsp90: FKBP51, FKBP52, PP5, and CHIP [
19]. By forming complexes with Hsp90, FKBP51 can influence the responsiveness of these receptors to steroid hormones, which are pivotal in regulating immune responses, inflammation, and cellular stress [
37]. FKBP51 modulates the conformation and activity of specific client proteins like the glucocorticoid receptor (GR) and regulates various kinase cascades including insulin signaling [
37]. Comorbidities like diabetes are recognized to exacerbate Aβ plaques in the context of age-related disorders. Inhibiting insulin binding impairs the brain’s synaptic plasticity, neuroinflammation, and the brain’s ability to metabolize glucose. A decrease in synaptic plasticity in AD individuals is one of the more noticeable ailments since it represents cognitive decline. Also, as a result of a decrease in synaptic plasticity, neuroinflammation activates and promotes tau accumulation [
38]. Increased levels of FKBP51 in conjunction with age and stress are seen to rise in individuals with AD [
19]. In transgenic mice models, LA1011 was shown to disturb the binding of FKBP51 to allow for Hsp90 to reduce disease-causing agents [
19]. The results of the study indicate that there is competition for binding to the hydrophobic pocket at the C-terminal end of Hsp90 between the helical extension of FKBP51’s TPR domain and LA1011 [
19]
Figure 1. The dihydropyridine LA1011 complex was also shown to decrease tau protein aggregation and Aβ plaque aggregation possibly via Hsp90 interactions in AD mice [
17,
19].
2.2. Temperature on Hsp90 Activity
Extremely high temperatures can induce cellular stress, which in turn can affect the expression and activity of HSP90 and other heat shock proteins. These proteins help cells cope with stress by preventing protein damage and aggregation. Observations revealed that sleep deprivation led to a rise in core body temperature, suggesting a connection between sleep deprivation and Aβ production in AD [
39]. Inflammatory environments in individuals with AD create a domino effect in the Hsps’ response to antagonistic environments; inflammation downregulates Hsp production since the cell is incapable of managing the stress, increasing Aβ aggregates [
38]. In disease conditions, Hsp90 has been shown to play a key role in aggregated protein processing and homeostasis. Slight changes in temperature from 37 °C to 39 °C led to enhanced Aβ40 and Aβ42 production in cell culture experiments [
39]. Hsp90 knockdown experiments at higher temperatures showed increased γ-secretase complex formation and the abolishment of increased Aβ production. Mouse model studies conducted at standard and higher temperatures showed that higher temperatures resulted in increased Hsp90, PS1-CTF, NCT, and γ-secretase complex levels.
2.3. Adenosine Triphosphate (ATP)-Mediated Changes in Hsp90 Activity
Neurons’ proper function relies on the efficient production of energy by the mitochondria. Mitochondrial dysfunction leads to reduced ATP generation, impairing neuronal activity and contributing to the cognitive decline observed in AD. Mitochondrial dysfunction is a significant contributor to the pathogenesis of AD and other neurodegenerative diseases, producing higher levels of reactive oxygen species (ROS) and NO, leading to oxidative stress [
40,
41]. Elevated ROS levels can cause damage to mitochondrial components, including lipids, proteins, and DNA, expediting impaired mitochondrial function. This damage can disrupt the electron transport chain, reduce ATP production, and compromise mitochondrial integrity, contributing to a vicious cycle of increasing ROS production and worsening mitochondrial dysfunction. Enhancing mitochondrial health could alleviate several aspects of AD progression and provide a multidimensional strategy to address the complex nature of the disease [
42,
43,
44]. The Hsp90 chaperone machinery is an ATP-dependent process. Essentially, ATP prevents Hsp90 chaperones from performing their inhibitory mechanisms, indicating that chaperones can be manipulated [
45,
46]. Hsp90 undergoes conformational changes as it interacts with client proteins. These changes are fundamental for the proper folding and stabilization of client proteins [
47,
48]. ATP hydrolysis provides the energy needed to facilitate these conformational changes. According to a recent study, ATP reduces Hsp90′s inhibitory impact on Aβ40 fibrillation by decreasing the hydrophobic surface of Hsp90 [
45]. When Hsp90 proteins are present, there is no indication of secondary β-sheet structure conformation, and the signal associated with the initial secondary Aβ40 random coil structure gradually diminishes over time, ultimately disappearing as incubation continues possibly due to Hsp90 and Aβ40 large aggregates forming (
Figure 1) [
45]. These findings suggest that Hsp90′s presence keeps Aβ mostly in a monomeric form. Further studies are needed to confirm the effect Hsp90 and ATP have on Aβ fibrillization in AD and other neurodegenerative diseases.
Figure 1. Schematic illustration of recent reports highlighting the importance of HSP90 in Aβ processing.
3. BRICHOS Chaperone Domain
3.1. BRICHOS Interactions
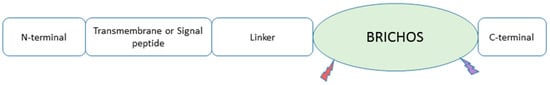
A chaperone that is due for major discussion is the BRICHOS chaperone domain. As shown in
Figure 2, all BRICHOS-containing proteins have an N-terminal cytosolic segment, a hydrophobic transmembrane or a signal peptide region, a linker segment, and the BRICHOS domain. The C-terminal segment is present in all BRICHOS except proSP-C [
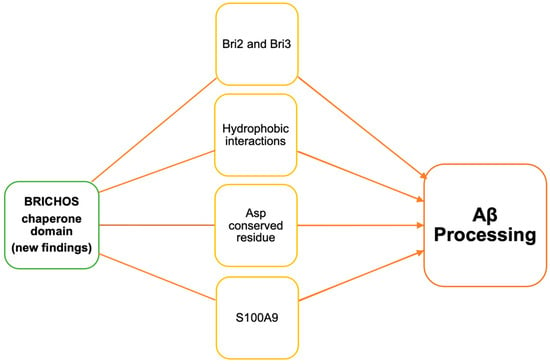
21]. Recent studies have shown this chaperone’s involvement in Aβ processing in various ways (
Figure 3). Österlund et al. (2022) used the structure prediction algorithm AlphaFold2 and mass spectrometry (MS) to define the BRICHOS domain’s interaction with Aβ. They used the BRICHOS-domain containing proform of lung surfactant protein C (proSP-C) for X-ray crystallography [
49]. This proSP-C protein is reported to inhibit AB fibrilization [
50]. Furthermore, Shimozawa et al. showed that BRICHOS interacts with multiple proteins [
51]. They used mice brain slice cultures incubated with recombinant BRICHOS Bri2 to perform proteomic analysis. They detected the binding of the Spectrin alpha chain, Spectrin beta chain, Myosin-10, unconventional Myosin-Va, Drebrin, Tubulin beta-2A chain, and Actin cytoplasmic 1 to BRICHOS Bri2.
Figure 2. A model illustrating the BRICHOS domain-containing protein. It contains an N-terminal cytosolic segment, a transmembrane or a signal peptide region, a linker segment, the BRICHOS domain, and a C-terminal segment. Mutations in Bri2 BRICHOS are shown as tiny lightning bolt shapes; red represents the D148N mutation and purple represents the R221E mutation.
Figure 3. Schematic illustration of recent reports on BRICHOS domain-containing proteins involved in Aβ processing.
3.2. S100A9 and Bri2 BRICHOS
Another recent study by Manchanda et al. (2023) generated a mutant form of recombinant human Bri2 BRICHOS R221E and tested it in AD animal models [
52]. Their findings suggest that the mutant form is more efficient than the wild-type form in preventing Aβ42-mediated toxicity in mouse slice culture. Furthermore, using the mouse model, they also showed that the mutated recombinant form is BBB-permeable. They also performed behavioral testing after treating an AD mouse model with the mutated Bri2 BRICHOS when AD pathology began to be observable through symptoms. The behavioral testing data showed that mutated Bri2 BRICHOS helped in boosting recognition and working memory, as was observed through an object recognition test. It is important to note that these positive results were only seen if the treatment started as soon as AD pathology began because this treatment did not show any improved symptoms if it was delayed by 4 months. To confirm these results, the amount of mutated Bri2 BRICHOS found in the brain correlated with the improvement and regaining of recognition and memory [
52].
Andrade-Talavera et al. investigated the effect of human recombinant Bri2 BRICHOS on S100A9 amyloid kinetics [
53]. S100A9 was reported to contribute to amyloid processing and neuroinflammation in AD, Parkinson’s disease, and traumatic brain injury. It forms an intracellular amyloid oligomer and Aβ co-aggregates in AD [
54,
55,
56]. S100A9 is an amyloidogenic polypeptide that is inhibited by recombinant Bri2 BRICHOS. The BRICHOS chaperone is seen to cap amyloid fibrils, which not only reduces aggregation but also decreases the inflammatory response (
Figure 3) [
53].
3.3. BRICHOS Domain Dependent on Conserved Asp Residue
While all of these recent developments are noteworthy and important, Chen et al.’s data on the conserved aspartate (D) residue in the BRICHOS domain provides further evidence of the potential of this chaperone [
57]. This particular residue has been consistent across most BRICHOS domain-containing proteins. A few BRICHOS domain-containing proteins have an aspartate to asparagine (N) mutation. Human Bri2 BRICHOS D148N mutation promotes dimerization without affecting function. Their findings suggest that the conserved aspartate residue’s pKa value ranges from pH 6.0 to 7.0, and due to its ionized state, aspartate promotes the domain’s structural flexibility (
Figure 3) [
57]. This chaperone’s role in the context of Hsp90 in Aβ processing is yet to be defined. Hence, future studies will help understand the role of the Bri2 BRICHOS chaperone in AD and its therapeutic potential.
3.4. Bri3 BRICHOS Domain Chaperone’s Efficiency
Poska et al. compared the efficiency of BRICHOS Bri2 and Bri3 against Aβ fibrillar and non-fibrillar aggregation [
23]. Bri2 and Bri3 BRICHOS domains have approximately 60% identical sequences, which indicates similarities in their structure and functions. Bri2 and Bri3 involvement in Aβ has been previously reported [
58,
59]. A recent report from Poska et al. suggests that Bri2 and Bri3 BRICHOS have differences in regulating Aβ. Bri2 was found to be more potent against Aβ fibrillar aggregation whereas Bri3 was more potent against non-fibrillar aggregation (
Figure 3). Bri3 is only expressed in the CNS whereas Bri2 is expressed in the CNS as well as peripheral tissues [
60,
61]. Overall, Bri2 and Bri3 have different tissue distributions and capacities as molecular chaperones. Future studies using mutants and AD models will help in fully understanding their involvement in AD pathogenesis.