The CD133 cell membrane glycoprotein, also termed prominin-1, is expressed on some of the tumor cells of both solid and blood malignancies. The CD133-positive tumor cells were shown to exhibit higher proliferative activity, greater chemo- and radioresistance, and enhanced tumorigenicity compared to their CD133-negative counterparts. The CD133-positive cells are related to the cancer stem cell subpopulation in many types of cancer. For this reason, CD133 is regarded as a potential prognostic biomarker in oncology.
- CD133
- prominin-1
- cancer biomarker
- cancer
1. Introduction
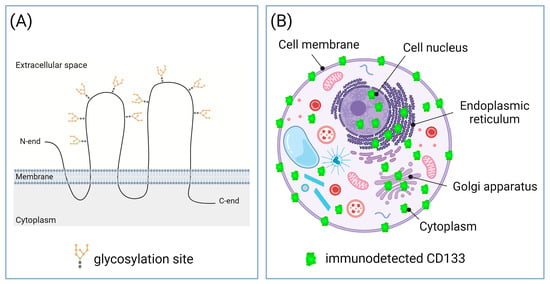
2. Putative Explanation of the Cause-and-Effect Relationship between CD133 Expression and More Malignant Phenotype of Tumor Cells
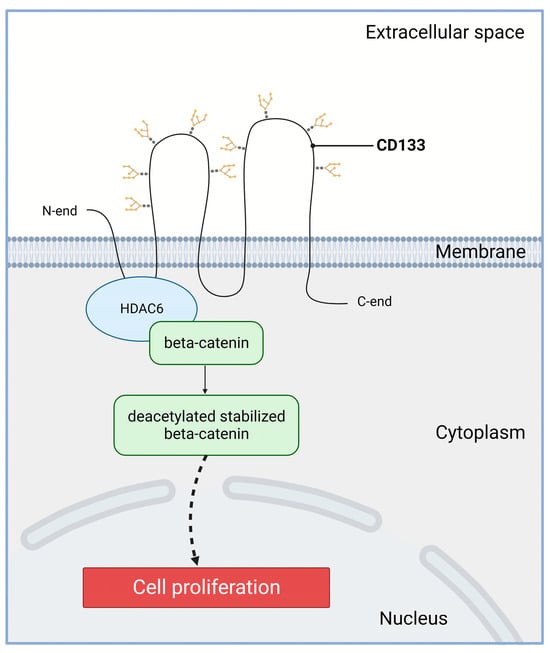
2.1. Increased Proliferation
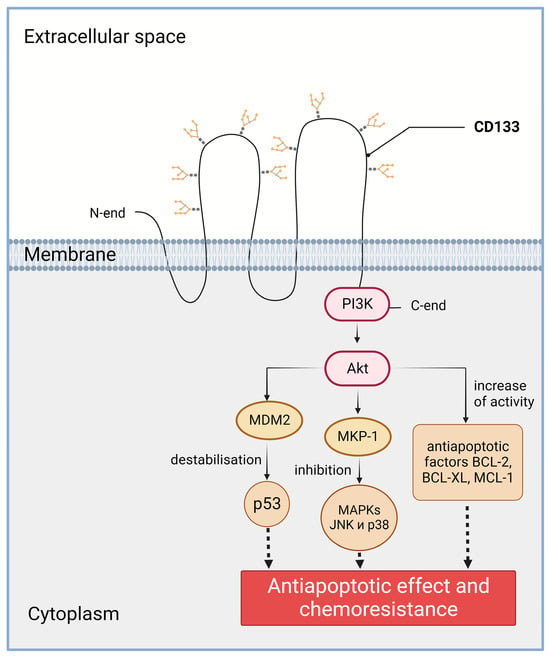
2.2. Increased Chemo- and Radioresistance
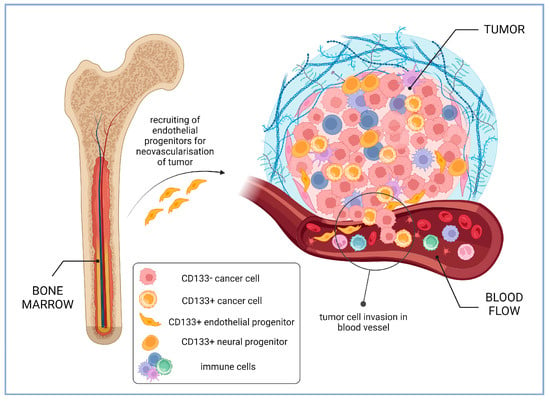
2.3. Increased Invasion and Metastasis
2.4. CD133 as a Target of Therapies Aimed to Eliminate the Most Malignant Cells
3. Association of CD133 with Cancer Progression and Poor Prognosis
For some tumors, including renal cell carcinoma and esophageal cancer, a correlation between CD133 expression and the unfavorable disease course was not found even in meta-analyses [227,228].
4. Possible Reasons for the Discrepancies in the Data on the Association of CD133 with the Disease Severity
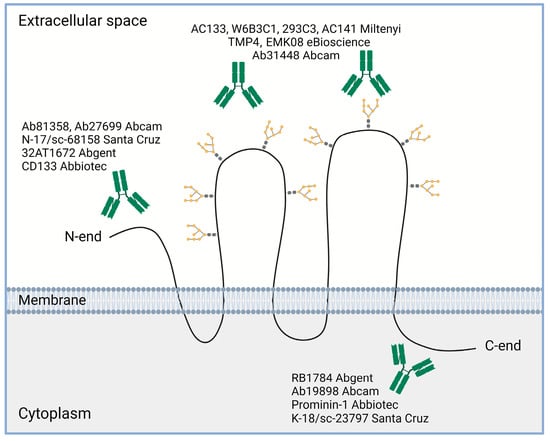
4.1. Different CD133 Immunodetection Techniques
4.2. Some CD133-Positive Cells Present within Tumors Are Normal, Benign Cells
4.3. Subcellular Localization of CD133 May Vary at Different Stages of Carcinogenesis
5. Conclusions
209. Xu, N.; Kang, Y.; Wang, W.; Zhou, J. The Prognostic Role of CD133 Expression in Patients with Osteosarcoma. Clin. Exp. Med. 2020, 20, 261–267, doi:10.1007/s10238-020-00607-6.
210. Han, M.; Guo, L.; Zhang, Y.; Huang, B.; Chen, A.; Chen, W.; Liu, X.; Sun, S.; Wang, K.; Liu, A.; et al. Clinicopathological and Prognostic Significance of CD133 in Glioma Patients: A Meta-Analysis. Mol. Neurobiol. 2016, 53, 720–727, doi:10.1007/s12035-014-9018-9.
211. Zhao, Y.; Peng, J.; Zhang, E.; Jiang, N.; Li, J.; Zhang, Q.; Zhang, X.; Niu, Y. CD133 Expression May Be Useful as a Prognostic Indicator in Colorectal Cancer, a Tool for Optimizing Therapy and Supportive Evidence for the Cancer Stem Cell Hypothesis: A Meta-Analysis. Oncotarget 2016, 7, 10023–10036, doi:10.18632/oncotarget.7054.
212. Fan, Z.; Li, M.; Chen, X.; Wang, J.; Liang, X.; Wang, H.; Wang, Z.; Cheng, B.; Xia, J. Prognostic Value of Cancer Stem Cell Markers in Head and Neck Squamous Cell Carcinoma: A Meta-Analysis. Sci. Rep. 2017, 7, 43008, doi:10.1038/srep43008.
213. Zhong, C.; Wu, J.-D.; Fang, M.-M.; Pu, L.-Y. Clinicopathological Significance and Prognostic Value of the Expression of the Cancer Stem Cell Marker CD133 in Hepatocellular Carcinoma: A Meta-Analysis. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2015, 36, 7623–7630, doi:10.1007/s13277-015-3487-y.
214. Chen, E.; Zeng, Z.; Bai, B.; Zhu, J.; Song, Z. The Prognostic Value of CSCs Biomarker CD133 in NSCLC: A Meta-Analysis. Oncotarget 2016, 7, 56526–56539, doi:10.18632/oncotarget.10964.
215. Tao, Y.; Li, H.; Huang, R.; Mo, D.; Zeng, T.; Fang, M.; Li, M. Clinicopathological and Prognostic Significance of Cancer Stem Cell Markers in Ovarian Cancer Patients: Evidence from 52 Studies. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 1716–1726, doi:10.1159/000489586.
216. Li, X.; Zhao, H.; Gu, J.; Zheng, L. Prognostic Value of Cancer Stem Cell Marker CD133 Expression in Pancreatic Ductal Adenocarcinoma (PDAC): A Systematic Review and Meta-Analysis. Int. J. Clin. Exp. Pathol. 2015, 8, 12084–12092.
217. Yiming, L.; Yunshan, G.; Bo, M.; Yu, Z.; Tao, W.; Gengfang, L.; Dexian, F.; Shiqian, C.; Jianli, J.; Juan, T.; et al. CD133 Overexpression Correlates with Clinicopathological Features of Gastric Cancer Patients and Its Impact on Survival: A Systematic Review and Meta-Analysis. Oncotarget 2015, 6, 42019–42027.
218. Li, Z.; Yin, S.; Zhang, L.; Liu, W.; Chen, B.; Xing, H. Clinicopathological Characteristics and Prognostic Value of Cancer Stem Cell Marker CD133 in Breast Cancer: A Meta-Analysis. OncoTargets Ther. 2017, 10, 859–870, doi:10.2147/OTT.S124733.
227. Cheng, B.; Yang, G.; Jiang, R.; Cheng, Y.; Yang, H.; Pei, L.; Qiu, X. Cancer Stem Cell Markers Predict a Poor Prognosis in Renal Cell Carcinoma: A Meta-Analysis. Oncotarget 2016, 7, 65862–65875, doi:10.18632/oncotarget.11672.
228. Kamil Mohammed Al-Mosawi, A.; Cheshomi, H.; Hosseinzadeh, A.; M Matin, M. Prognostic and Clinical Value of CD44 and CD133 in Esophageal Cancer: A Systematic Review and Meta-Analysis. Iran. J. Allergy Asthma Immunol. 2020, 19, 105–116, doi:10.18502/ijaai.v19i2.2756.
This entry is adapted from the peer-reviewed paper 10.3390/ijms242417398